Imagen de portada: Rock Islands Southern Lagoon (Koror). Palau. [Fuente: tripadvisor]
I jumped in the river and what did I see?
Black-eyed angels swam with me
[Pyramid song. Radiohead, 2011]
Las pirámides del antiguo Egipto se construyeron entre el 2700 y el 1500 A.C.
Y entre todas ellas, la Gran Pirámide de Giza es la única de las 7 maravillas de la Antigüedad que continúa todavía en pie.
La gran pirámide de Giza, también conocida como de Keops. Autora: Nina Aldin Thune. Fuente: Wikimedia Commons.
Como bien sabemos se trataba de tumbas monumentales para los faraones (jefes del estado y líderes religiosos al mismo tiempo). Por encima de faraón no se ha inventado nada ¿verdad?
En su vasta mayoría se trató de hombres, aunque algunas mujeres también ocuparon el cargo como Hatshepsut y Nefertiti. Sin olvidar por supuesto a Cleopatra VII, última reina de la dinastía Ptolemaica hasta que Roma conquistó Egipto (en el 30 A.C.) y lo anexionó como provincia del Imperio. ¿Les dirían que eran «de provincias» al visitar Roma? vaya usted a saber…
Las pirámides han despertado todo tipo de teorías e hipótesis loquísimas como la participación de extraterrestres en su construcción (¿os suena un tal Elon Musk? pues en 2020 dijo «Aliens built the pyramids, obv»). O que las más famosas, las de Giza, se alinean con Mercurio, Venus y Saturno. Incluso que incluyen jeroglíficos con medios de transporte modernos, etc. Lo que sí es cierto es que su majestuosidad y aureola de misterio (aún desconocemos con exactitud cómo se construyeron) ha cautivado durante siglos a la humanidad.
Las pirámides contemplan el paso del tiempo en medio de un paisaje árido al oeste del Nilo. Yo nunca he estado allí pero las imagino rodeadas de tierra y arena, expuestas a un calor sofocante.
Por eso me fascinó descubrir que existe un dinoflagelado fotosintético tropical que vive en la arena (sand-dwelling en inglés) y que se llama Pyramidodinium.
Ya imaginaréis a qué se parece…
Así es Pyramidodinium atrofuscum. Las formas piramidales (1, 2, 3 y 4), un quiste (5) y la forma móvil flagelada (6). Autores: Horiguchi y Sukigara (2005). Fuente: wiley.com
Lo encontraron unos investigadores japoneses en el archipiélago de Palau y sus razones para denominarlo así fueron las siguientes (traduzco del inglés):
«Pyramidodinium atrofuscum (género y especie nueva). Etimología: del griego «pyramis» que indica la forma de la célula inmóvil vegetativa (en forma de pirámide) y el latín «atrofuscum» (marrón negruzco) por el color del citoplasma.»
[Horiguchi y Sukigara, 2005]
Su aspecto a mí me recuerda a un juego de niños, el «comecocos» de papel. De hecho, las paredes del dinoflagelado son también de celulosa.
Pero vais a alucinar todavía más. Al dinoflagelado pirámide lo aislaron en un lago marino llamado «El Lago de las Medusas». Un lugar mágico y digno de «La Isla Misteriosa» de Jules Verne.
Dicho lago está conectado con el océano a través de fisuras y túneles superficiales en lo que era un antiguo arrecife. Sus condiciones ambientales (clima tropical, lluvias copiosas, rodeado por vegetación y paredes rocosas que frenan el viento) provocan una estratificación permanente. Su capa profunda comienza a partir de 15 metros, no tiene oxígeno y es tóxica para el ser humano por la acumulación de sulfuro de hidrógeno. Sobre ella (y gracias a ese H₂S) existe una capa fina de bacterias púrpuras fotosintéticas del azufre (como las que tiñen de rosa en verano la playa do Adro, en Vigo).
El Lago de las Medusas, en la isla Eil Malk del archipiélago de Palau. Los puntos amarillos que se ven en el agua son medusas. Autor: Lukas. Fuente: Wikipedia
En la capa superior oxigenada es donde proliferan de forma permanente las medusas que le dan nombre al lago. Hay 2 especies y una de ellas es dorada (Mastigias papua) porque contiene dinoflagelados simbiontes. Los suyos pertenecen al género Cladocopium (hasta hace poco se les llamaba Symbiodinium «clado C»). Las medusas se mueven al ritmo del sol y no suponen ningún riesgo para la salud, así que puedes nadar entre ellas (con un guía local) aunque el buceo está completamente prohibido.
Las medusas doradas (Mastigias papua) en el Lago de las Medusas. Fuente: supercurioso.com
A Pyramidodiniumlo aislaron en una zona arenosa y poco profunda del lago. No le gusta moverse así que pasa la mayor parte del tiempo en el fondo en forma de pirámide.
De vez en cuando se divide y salen por el pico de la pirámide dos células móviles con aspecto de dinoflagelado desnudo (como Gymnodinium) que nadan durante un rato.
Esa transformación se observa en los cultivos de laboratorio entre 3 y 6 horas después de encender la luz. Las células móviles son delicadas y a la mínima cesan de nadar y vuelven al fondo. Primero se vuelven redondas y luego cuadradas hasta convertirse de nuevo en pirámides. Está claro que prefieren la vida contemplativa. Permanecen así, como el faraón dentro de su tumba unos 5 días antes de dividirse y dar otro paseo.
En 2017 se descubrió una segunda especie en este género: Pyramidodinium spinulosum. Fue el mismo grupo de japoneses y esta vez la encontraron casi a 4o metros de profundidad, en los fondos arenosos de la isla de Mageshima (Pacífico subtropical, Japón). Aunque en este caso en vez de pirámides las células inmóviles parecen lentejas espinosas.
Pyramidodinum spinulosum. Fuente: Horiguchi y col. (2017).
Qué queréis que os diga, me quedo con la primera especie…
Esa que vive en un lago tropical donde reinan las medusas sobre miles de pirámides en la arena.
Referencias:
Hamner WM y col. The Physical, chemical, and biological characteristics of a stratified, saline, sulfide lake in Palau. Limnol. Oceanogr. 27(5): 896–909. (1982).
Horiguchi T. & Sukigara C. Pyramidodinium atrofuscum gen. et sp. nov. (Dinophyceae), a new marine sand-dwelling coccoid dinoflagellate from tropical waters. Phycol. Res. 53(4): 247-254. (2006).
Horiguchi T. y col. Pyramidodinium spinulosum sp. nov. (Dinophyceae), a sand-dwelling non-motile dinoflagellate from the seafloor (36 m depp) off Mageshima Island, Kagoshima, Japan. Phycol. Res. 65(4): 272-279. (2017).
Vega de Luna F y col. Photosynthetic capacity of the endosymbiotic dinoflagellate Cladocopium sp. is preserved during digestion of its jellyfish host Mastigias papua by the anemone Entacmaea medusivora. FEMS Microbiol Ecol. 95(10):fiz141. (2019).
https://fitopasion.com/wp-content/uploads/2025/02/palau-caroline-islands.jpg563900Francisco Rodriguezhttps://fitopasion.com/wp-content/uploads/2021/06/logo-FITOPASION.pngFrancisco Rodriguez2025-02-10 21:01:242025-02-10 21:01:24Pirámides bajo el mar
Imagen de portada: playa de Mogor. Autor: F. Rodríguez
El dinoflagelado Lepidodinium es responsable del verde que tiñe las Rías Baixas en las últimas semanas. Probablemente sea Lepidodinium chlorophorum. Ya veremos…
En una entrada reciente hablé sobre él, pero no esperaba encontrármelo en Galicia y mucho menos con una proliferación tiñendo el mar en la costa y varias rías entre Vigo y Finisterre.
Aquí podéis verlo en una muestra natural del puerto de Finisterre el 29 de agosto de 2024.
Las mal llamadas mareas rojas (proliferaciones de microalgas que colorean la superficie del mar pero que ni son mareas ni en muchos casos rojas), son habituales en Galicia. Suelen ser marrones, naranjas, granates o rojas en función de los pigmentos de las células y de su abundancia. En extrañas ocasiones son verdes, como esta.
Las mareas rojas son un fenómeno natural que se conoce desde antiguo relacionado con la elevada productividad debida al afloramiento estacional y los aportes de aguas continentales, entre otros factores. Las microalgas no necesitan de contaminación para proliferar y teñir el mar. Como regla general, cuando superan 1 millón de células por litro su color es visible a simple vista.
En Galicia no podemos achacar las mareas rojas a la acción humana -sin pruebas fehacientes- porque la hipótesis más común, sencilla y probable es la de un suceso natural.
Estas son algunas imágenes de otros autores que comparto con su permiso…
El pase de diapositivas requiere JavaScript.
La época de crecimiento del fitoplancton se extiende entre primavera y otoño en las rías gallegas. Y en ese periodo se producen mareas rojas asociadas al desarrollo del fitoplancton (y zooplancton), en condiciones climatólogicas y oceanográficas favorables. Esas mareas rojas también se deben a una combinación de factores biológicos y ecológicos de los propios organismos que les permite mantener poblaciones densas y competir mejor por los recursos (luz y nutrientes).
Entonces, nada nuevo bajo el mar. ¿O sí?
PUES SÍ
La mayoría de mareas rojas en Galicia las producen dinoflagelados: casi 2 de cada 3 basándonos en el registro –cualitativo a partir de observaciones en artículos científicos y prensa-, de 112 mareas rojas que publicamos en una reciente revisión, la primera en Galicia (Rodríguez y col. 2024).
Os muestro una gráfica de ese trabajo con los grupos del fitoplancton, especies responsables y colores aproximados de las mareas rojas entre 1916 y 2023 en Galicia. Encontrar las verdes es como jugar a ¿dónde está Wally?. Entre ellas veréis que solo hay una de Lepidodinium.
Esa marea verde sucedió entre julio y septiembre de 1989 en las Rías de Vigo (Reguera y col. 1995) y Pontevedra (frente a Marín, cerca de la isla de Tambo; Jiménez y col. 1992). Por entonces apenas se hacían análisis genéticos en estudios de fitoplancton y tampoco existía el conocimiento actual sobre dinoflagelados como aquel.
Se identificó como Gyrodinium aureolum o Gyrodinium sp., pero la descripción del mismo y la coloración del mar («bright and green patch» y «emerald green» según esos autores) encajan con una proliferación de Lepidodinium que llegó en Pontevedra a los 70 millones de células por litro.
Y es que no existe otro dinoflagelado que produzca ese color verde brillante «esmeralda». Lo único que no me cuadra es que los resultados de pigmentos no citan a la clorofila b, que debería ser muy abundante porque es propia de cloroplastos verdes como los de Lepidodinium.
Canido, desde el puente de Toralla. 17 de septiembre 2024. Autor: F. Rodríguez
En entradas anteriores hablé sobre Lepidodinium (1,2) pero dejaré aquí un resumen para hacerle la ficha a nuestro protagonista de hoy:
Es un dinoflagelado desnudo, sin cubierta de celulosa o calcárea en sus células. El género Lepidodinium incluye a día de hoy 2 especies indiferenciables al microscopio óptico: L. chlorophorum y L. viride. Para distinguirlas hay que usar genética y microscopía electrónica. Lepidodinium viride posee unas escamitas diminutas ausentes en L. chlorophorum. La especie descrita en el Atlántico es L. chlorophorum (salvo contadas excepciones) así que es probable que se trate de la responsable de la «marea verde» en Galicia.
No es tóxico ni produce bioluminiscencia. Soporta un amplio rango de salinidad y temperatura. Sus proliferaciones suelen asociarse con estratificación térmica y aportes de aguas fluviales. No supone ningún peligro para el baño, pero como siempre ante cualquier mancha desconocida en el agua es aconsejable tener precaución.
Su color verde se debe a cloroplastosadquiridos de microalgas verdes (pedinofíceas). Esto es una excepción entre los dinoflagelados ya que ningún otro posee cloroplastos permanentes de este tipo. Salvo en Lepidodinium, todos proceden de grupos de la línea evolutiva roja. Además, posee un pigmento (carotenoide) exclusivo: lepidoxantina. Con él podemos trazar su presencia y abundancia en el mar. Por cierto: dicho pigmento lo caracterizaron un grupo de investigadores de Vigo (Gavalás-Olea y col. 2016).
Lepidodinium libera grandes cantidades de sustancias poliméricas.Mucus para entendernos. Con ello puede alterar la viscosidad del agua y parece que podría jugar un papel importante en el mantenimiento de sus proliferaciones, formando un microambiente favorable para el crecimiento de bacterias y el reciclado de nutrientes que aprovecharía el propio dinoflagelado.
Algunos blooms masivos de Lepidodinium se han asociado con muertes de peces e invertebrados, principalmente en la costa atlántica de Francia y Chile. En Francia desde los años 90′, especialmente en el sur de Bretaña, afectando a peces y moluscos. Y en Chile en verano de 2021 donde produjo la muerte de 162.000 salmones en la región de Los Lagos. Las causas no están del todo claras pero todo apunta al consumo de oxígeno cuando declinan sus poblaciones (y quizás también por el mucus que podría afectar a moluscos filtradores).
A todo esto añadiré mis observaciones personales en estos días. El color del agua es bonito y puede costar diferenciarlo de una idílica postal veraniega con aguas turquesas cuando no está muy concentrado el bloom.
El agua en la orilla de la playa de Mogor durante el bloom de Lepidodinium. 8 de septiembre 2024. Autor: F. Rodríguez
En las zonas donde se acumula, como el muelle de Aguete en la Ría de Pontevedra, se forman espumas y grumos en superficie con un fuerte olor que llega a ser nauseabundo. Ese olor fuerte «a mar» y que se ha descrito como a «col hervida» (o «berberecho» según mi chica) se debe a la liberación de un gas (dimetilsulfuro) por parte de las microalgas.
El bloom de Lepidodinium acumulado dentro del muelle del puerto de Aguete. 10 de septiembre 2024. Autor: F. Rodríguez
A mí ese olor me recuerda muchísimo al tufo que producían los gusanitos de Symsagittifera roscoffensis en nuestro laboratorio (llenos de microalgas verdes simbiontes del género Tetraselmis).
En el fondo arenoso se acumulan grumos verdes resbaladizos que se escapan entre las manos si los intentas coger. El oleaje los va rompiendo en fragmentos más pequeños y degradando hasta volverlos marrones y dejarlos en la orilla (La Voz de Galicia, 11-IX-2024)
Otro tema interesante es que el bloom parece concentrarse bajo la superficie y no arriba de todo a pleno sol como es habitual en otros dinoflagelados como Alexandrium. En el siguiente vídeo grabado desde el puente de Toralla (Ría de Vigo) podéis apreciar como el agua es más transparente en superficie y se vuelve verde al descender; así hasta el fondo (unos 4 metros).
La espuma resultante de los agregados de células de Lepidodinium durante el bloom se observa también en aguas alejadas de la orilla, a lo largo de frentes de marea y zonas de acumulación por acción de las corrientes. Esa espuma es verdosa-amarillenta, su aspecto también cambia según la iluminación.
Espumas procedentes del bloom de Lepidodinum. Playa de Mogor (Marín, Ría de Pontevedra). 8 de septiembre 2024. Autor: F. Rodríguez
Yo nunca había visto este tipo de espuma y cuando la observé en directo inmediatamente me vino a la cabeza una imagen que compartió La Voz de Galicia (15-VIII-2024) en la playa de Patos (Nigrán, Ría de Vigo).
Por aquellas fechas se produjo un cierre cautelar al baño en dicha playa; primero se creyó que eran macroalgas y luego se publicó que eran microalgas, que no había peligro y que se abría al baño la zona. Hablamos de la foto en el IEO y pensamos qué cosa tan rara. Pero habían pasado varios días y no fui a ver si quedaban rastros de aquello. Recordaba a las manchas de polen en primavera, a saber…
El 29 de agosto me envió Ana García (La Voz de Galicia) un vídeo de un marinero (Julio, del Fish-Terra) con el agua verde brillante en el puerto de Fisterra. Aquel día Ana presentaba su libro «Mar de Ardora» en Vimianzo al que me había invitado para participar con una breve charla.
Vimianzo está a media hora en coche de Fisterra y al finalizar el evento por la tarde (después de pensármelo dos veces porque el regreso a Vigo son 2 buenas horas de coche) tomé la afortunada de decisión de ir hasta allí. Llegué a las 21:30, casi de noche, pero me pareció apreciar que incluso así el agua estaba más oscura de lo normal. Cogí una muestra en una de las rampas del muelle.
Al día siguiente en el IEO identifiqué a Lepidodinium y no por esperado fue menos sorprendente. Un hecho interesante pero anecdótico que recogió también La Voz de Galicia («Fenómeno verde en el puerto de Fisterra», 7-IX-2024). Y luego me fui de vacaciones hasta mediados de septiembre.
El fin de semana anterior a mi regreso (8 de septiembre) me llegaron muchísimas notificaciones de avistamientos del «mar verde» en las Rías de Vigo, Pontevedra y Muros. Desde aguas abiertas como Cabo Udra a playas en Nigrán, Vigo, Bueu y Marín, entre otras. También en Cíes, Bascuas y Carnota. Ya no era anecdótico y la atención mediática ha sido importante en prensa escrita, radio y televisión.
Lepidodinium (puerto de Fisterra, 29 de agosto 2024). A 400 aumentos. Autor: F. Rodríguez
¿Cuál es la explicación de esta «marea verde»? Se trata de una época propicia para las proliferaciones de dinoflagelados y el verano en las Rías Baixas parece no dejarnos este año con temperaturas por encima de la media para agosto y todavía en septiembre, con predominio de vientos favorables al afloramiento.
Es necesario analizar las muestras del material recogido durante el bloom y estudiar las condiciones climatológicas y oceanográficas en estas últimas semanas para entender por qué ha sucedido y por qué es tan extenso. En cuanto a ¿por qué Lepidodinium?, esto es un reto aún mayor al no haber apenas precedentes y quizás no haya ninguna respuesta evidente.
Hasta entonces, dados los precedentes que citaba antes, la buena noticia es que no parece haber ocasionado daños a la fauna marina en general ni a especies de interés comercial ya sean peces o marisco. Todo ello a pesar de la extensión geográfica y duración del bloom que en si mismo es un hecho preocupante. La razón es que su aparición podría suponer que la presencia de Lepidodinium sea más habitual en el futuro. La suma de un nuevo actor entre las especies de fitoplancton potencialmente nocivas (y por tanto a vigilar), en las rías gallegas. ¡Esperemos que no sea así!
NOTA: Si queréis enviarme fotos para completar esta entrada ¡estais invitados! los datos de los avistamientos y fechas también son MUY importantes para trazar el inicio y extensión del bloom.
Agradecimientos: a todas y todos los que habéis compartido avisos e imágenes sobre este bloom verde tan extraordinario: Julio del «Fish-Terra», Ana, Montse, Pablo, Garci, Julio Valeiras, José Luis y mis compañeras del grupo VGOHAB del IEO de Vigo. También a través de redes sociales (Rogelio y una seguidora suya en X). Si olvido a alguien ¡lo añadiré!
Referencias:
Gavalás-Olea A. y col. 19,19’-diacyloxy signature: an atypical level of structural evolution in carotenoid pigments. Org. Lett. 18(18):4642-5. (2016).
Jiménez C. y col. Green mass aggregations of Gyrodinium cf. aureolum Hulburt in the Ria of Pontevedra (north-west Spain). J. Plankton Res. 14:705-720. (1992).
Kamikawa R. y col. Plastid Genome-Based Phylogeny Pinpointed the Origin of the Green-Colored Plastid in the Dinoflagellate Lepidodinium chlorophorum. Gen. Biol. Evol. 7: 1133–1140. (2015).
Roux P. y col. Spatio-temporal dynamics and biogeochemical properties of green seawater discolorations caused by the marine dinoflagellate Lepidodinium chlorophorum along southern Brittany coast. Est. Coast Sci. 275:107950. (2022).
Roux P. Propriétés écologiques des efflorescences de Lepidodinium chlorophorum: de
l’écophysiologie cellulaire à l’impact sur l’écosystème. Sciences de la Terre. Nantes Université,
Français. NNT:2022NANU4026. tel-03851976. pp. 310. (2022).
Reguera B. y col. Autoecology and some life history stages of Dinophysis acuta Ehrenberg. J. Plankton Res. 17:999-1005. (1995).
Rodríguez F. y col. Red tides in the Galician rías: historical overview, ecological impact, and future monitoring strategies. Environ Sci Process Impacts. 26(1):16-34. (2024).
Yang Q-Q. Production of dimethylsulfoniopropionate, dimethylsulfide and acrylic acid
from marine microalgae. J. Sea Res. 190:102299 (2022).
Web: Millonarias pérdidas económicas en importante salmonera chilena a causa de un fenómeno de algas nocivas (mispeces.com).
https://fitopasion.com/wp-content/uploads/2024/09/Portada-1-scaled.jpg19282560Francisco Rodriguezhttps://fitopasion.com/wp-content/uploads/2021/06/logo-FITOPASION.pngFrancisco Rodriguez2024-09-19 16:59:342024-09-27 11:03:06Marea verde en las Rías Baixas
Imagen de portada: veneno de basilisco. Fuente: aminoapps.com
El veneno de basilisco es una sustancia muy poderosa y tóxica que sólo tiene una cura conocida: lágrimas de fénix. Este veneno es tan potente que puede matar a una persona en cuestión de minutos, lo que hace que la persona quede somnolienta y con visión borrosa antes de morir.
Las biotoxinas marinas suponen un riesgo sanitario, de ahí la necesidad de controlar su presencia, sobre todo en marisco, para evitar intoxicaciones. Y ese control lo realizan las autoridades de cada país; en el caso de la UE con la legislación europea en la mano.
Esa legislación detalla los niveles permitidos de biotoxinas responsables de síndromes como el amnésico, diarreico y paralizante (ASP, DSP y PSP), además de los métodos oficiales que permiten análisis homogéneos en la UE. Las leyes europeas también hablan de ciguatoxinas, cuya presencia está prohibida en productos de la pesca, al igual que la venta de peces de familias como Tetraodontidae («peces globo», por culpa de las tetrodotoxinas).
Pero existen otra serie de compuestos y síndromes que hoy por hoy quedan fuera de la norma europea. Aquí juegan un papel importante los proyectos de investigación para estudiar qué otras toxinas y organismos las producen, así como su toxicidad y acumulación en productos del mar para evaluar riesgos y en su caso actualizar leyes.
Los síndromes asociados con biotoxinas marinas se abrevian con siglas.
Ya mencioné ASP, DSP, PSP, pero si revisamos publicaciones sobre este asunto como «Marine Biotoxins» de la FAO encontraremos siglas como AZP (azaspiracid shellfish poisoning; azaspirácidos, incluidos en la legislación europea), CFP (ciguatera fish poisoning) y NSP (neurotoxic shellfish poisoning; brevetoxinas). Hasta aquí todo correcto…
Pero en algunas publicaciones encontrareis citas de VSP (venerupin shellfish poisoning), asociado a la venerupina. Y os preguntaréis ¿qué diablos es la venerupina?
Pues bien. Puedo adelantaros que tras revisar trabajos y consultar fuentes diversas…¡no tengo respuesta!. Aquí os hago un resumen de la búsqueda y explicación a la que he llegado sobre la venerupina y el VSP.
Los hechos son los siguientes. En marzo de 1942 se intoxicaron 324 personas tras comer almejas «asari» o japónica como la conocemos en Galicia (Venerupis semidecussata), cerca del lago Hamana (en Japón ¡claro!). El saldo final fue de 114 fallecidos.
24-48 horas después de la comida, las personas intoxicadas sufrieron dolor abdominal, náuseas, vómitos, dolor de cabeza, sangrado en la nariz y otras zonas, petequia generalizada (puntitos planos bajo la piel a consecuencia del sangrado), delirio y muerte en casos agudos. Las intoxicaciones sucedieron tras ingerir al menos unas 40 almejas. Un par de platos como el siguiente serían suficientes…
Udon con almejas asari, acompañado con algas wakame. Fuente: Gigazine
En marzo de 1943 y 1949 ocurrieron intoxicaciones similares esta vez por ostras (Crassostrea gigas) con 71 y 6 fallecidos respectivamente. El porcentaje de mortalidad fue inferior gracias a un veloz diagnóstico y tratamiento. Akiba y Hattori publicaron un estudio en 1949 en el que consiguieron extraer la toxina y demostrar sus efectos en perros, gatos, conejos y ratones. En todos ellos observaron daños hepáticos y hemorragias.
La toxicidad era organotrópica (con afinidad por órganos particulares) y no neurotrópica como la mayoría de toxinas conocidas. Para denominar a este nuevo síndrome usaron el nombre de las almejas japónicas y le llamaron «Venerupin poisoning» y «venerupina» a la toxina. Todo ello sin evidencias del agente causante ni de la identificación de la sustancia.
A finales de los años 50′ sucedieron episodios con síntomas de intoxicación PSP por consumo de marisco en la laguna de Óbidos (Portugal), relacionados con blooms del dinoflagelado Prorocentrum minimum. Luego, varios trabajos de Nakajima en el lago Hamana en los 60′ establecieron una relación entre la aparición de P. minimum en el agua y la toxicidad del marisco por venerupina o VSP.
Entre 1968-1971 se registraron nuevas intoxicaciones compatibles con VSP por consumo de almejas en la bahía de Okpo (isla Geoje, Corea del Sur). Sumaron un total 319 personas afectadas con 21 fallecidos. Sin pistas sobre el organismo responsable, la construcción de un astillero industrial en la zona «solucionó» años después el problema.
Con el paso del tiempo, nuevos estudios pusieron en entredicho las conclusiones de Nakajima y otros trabajos sobre la venerupina y la toxicidad de P. minimum, convirtiendo al VSP y a la venerupina en una incógnita sin solución. Y en resumen esto es lo que hay.
Si buscan la estructura de la venerupina o evidencias de su existencia no encontrarán nada.
Lo cierto es que desde los años 50′ y en décadas siguientes las proliferaciones y mareas rojas de P. minimum se han relacionado con toxicidad en marisco o intoxicaciones en personas y daños a la fauna marina en una larga lista de países como Angola, Noruega, Francia, China, Pakistán o EEUU. Y de todas ellas, solo las intoxicaciones de Noruegaen 1979tuvieron características similares al VSP de Japón. Pero no se avanzó más en el asunto.
Además, la mayoría de cepas aisladas de P. minimum no son tóxicas, excepto varias aisladas en 1989-90 en el Mediterráneo francés. Y en las francesas se detectaron compuestos neurotóxicos desconocidos que no provocaban los síntomas hepatotóxicos del VSP.
En otras zonas del mundo como Galveston (Texas, EEUU) cuando las ostras se alimentan durante blooms de P. minimum lo único que les pasa es que se vuelven rosadas. Esto no les produce daños ni toxicidad pero pierden atractivo y valor en el mercado. En 1999 un estudio taxonómico determinó que P. minimum y P. cordatum eran sinónimos y que el nombre válido para la especie sería P. cordatum.
El último capítulo sobre la posible identidad de la venerupina procede de Grecia. En 2012 se registró una toxicidad atípica en bioensayo de ratón y la presencia de tetrodotoxinas en mejillones en Rodopi (Grecia), coincidiendo con blooms de P. minimum (=P. cordatum). La hipótesis fue que la causa y que el misterioso síndrome VSP achacado a esta especie en el pasado podría deberse a tetrodotoxinas.
Mejillones griegos (M. galloprovincialis) preparados con la receta Saganaki. Fuente: mygreekdish.com
Pero faltaban muestras de P. cordatum para avanzar en esta idea. Después, Rodríguez y col. (2017) aportaron nuevos resultados al descubrir bacterias productoras de tetrodotoxinas en aquellos mejillones griegos, e identificar análogos de tetrodotoxinas y bacterias relacionadas con su producción en cultivos de P. cordatum. Así pues, la simbiosis entre bacterias y P. cordatum podría explicar esa toxicidad.
Dado el tiempo transcurrido y los datos disponibles sobre VSP quizás nunca podamos confirmar a ciencia cierta qué sucedió. Los síntomas de intoxicación por tetrodotoxinas coinciden en parte (náuseas, diarrea, vómitos, dolor abdominal), pero al ser neurotoxinas sus efectos se parecen más al PSP que a los descritos en 1942 en el lago Hamana. Para estar seguros necesitaríamos toxinas en marisco y microalgas de la misma zona; y que produjesen síntomas similaresal VSP.
En conclusión, y tras este viaje en la bibliografía, cada vez que lea venerupina pensaré en toxinas fantásticas como el veneno de basilisco de la saga de Harry Potter. La diferencia es que aquella intoxicación fue real y ello explicaría que la literatura científica siga citando a la venerupina. Después de todo –aunque no lo parezca– quizás sí exista…
Un basilisco comportándose como tal. Fuente: aminoapps.com
Referencias
Akiba T. & Hattori Y. Food poisoning caused by eating asari and oyster-toxic substance, venerupin. Jpn. J. Exp. Med. 20:271–284. (1949).
Grzebyk D. y col. Evidence of a new toxin in the red-tide dinoflagellate Prorocentrum minimum. J. Plankton Res. 19:1111–1124. (1997).
Heil C.A. y col. Prorocentrum minimum (Pavillard) Schiller: A review of a harmful algal bloom species of growing worldwide importance. Harmful Algae 4:449–470. (2005).
Hong S.S. y col. Studies on venerupin shellfish poison in Korea. Kor. J. Gastroenterol. 7:151-164. (1975).
Kawabata T. Part II. Fish-Borne Food Poisoning in Japan. G. Borgstrom (Ed.), Fish as Food, Vol. 2, Academic Press, London, p. 467-479. (1962).
Rodríguez I., y col. The association of bacterial C9-based TTX-like compounds with Prorocentrum minimum opens new uncertainties about shellfish seafood safety. Sci Rep. 20;7:40880. (2017).
Velikova V. & Larsen J. The Prorocentrum cordatum/Prorocentrum minimum taxonomic problem, Grana, 38:2-3, 108-112 (1999).
Vlamis A. y col. First detection of tetrodotoxin in greek shellfish by UPLC-MS/MS potentially linked to the presence of the dinoflagellate Prorocentrum minimum. Toxins 7:1779–1807. (2015).
https://fitopasion.com/wp-content/uploads/2022/12/venenobasilisco.jpg570570Francisco Rodriguezhttps://fitopasion.com/wp-content/uploads/2021/06/logo-FITOPASION.pngFrancisco Rodriguez2022-12-12 17:11:172023-12-17 22:21:36Venerupina y veneno de basilisco
Imagen de portada: mar de ardora en la ría de Vigo. Autor: Buceo Islas Cíes.
En la última semana se han publicado muchísimas imágenes de mareas rojas en las Rías Baixas, sobre todo en las de Vigo y Pontevedra, asociadas con bioluminiscencia («mar de ardora» le decimos por aquí). El aspecto de las manchas es el típico de Noctiluca. Su tamaño (cerca de 1 milímetro) permite observarlas a simple vista y si llenas una botella verás miles de bolitas flotando en una capa anaranjada (parecen aceite).
Ardora en la Ría de Vigo, 03-09-21 (Buceo Islas Cíes)
Areamilla, 05-09-2021 (Cangas do Morrazo). Autor: F. Rodríguez
De ellas he escrito numerosas veces en estos 11 años (ay josús…). Si escriben «Noctiluca» en el buscador del blog descubrirán entradas como esta o esta otra con más información sobre ellas. Hoy nos bastará con recordar que son dinoflagelados heterótrofos (no realizan fotosíntesis), que zampan gran variedad de plancton.
Dejemos las Rías Baixas y el «Vigocentrismo». En meses anteriores, durante julio y agosto, se publicaron numerosas noticias en prensa, televisión y RRSS sobre mares de ardora en A Costa da Morte («Un baño no mar de ardora coa Vía Láctea de compañeira en Corme«; GCIENCIA, 8-VII-2021; «¿Cuándo y por qué arde el mar de la Costa da Morte?»; La Voz de Galicia, 13-VII-2021).
En cuanto a los vídeos destacaría este enlace de @drewkorme entre el 15-16 de agosto en Ermida (Corme). En su cuenta de instagram descubrirán imágenes preciosas. Pero como no puedo enlazar su vídeo desde facebook aquí va otro de una playa cercana (Niñóns, Ponteceso), de Oscar Blanco.
Las Rías Altas de Galicia poseen un encanto natural único. Y zonas como Carnota y la Ría de Corme e Laxe parecen idóneas para disfrutar de la ardora durante el verano. Es sólo una impresión subjetiva porque no hay registros sistemáticos ni seguridad al 100% de que la vayas a ver. Sobre esto hablaremos al final…
Basta leer noticias como «Cinco playas gallegas para disfrutar del Mar de Ardora» (La Voz de Galicia, 1-IX-2021) para darse cuenta de que A Costa da Morte es un destino interesante para disfrutar de la ardora.
Además, alejada de grandes ciudades, dicha región posee mucha menos contaminación lumínica que las Rías Baixas o el área de influencia de A Coruña donde si no ponemos farolas cada 50 metros alumbrando al mar no nos quedamos tranquilos (no vaya a ser que salga un Kraken y no lo veamos).
Animado por estas noticias (y antes de que las Noctilucas invadiesen el sur de Galicia), nos fuimos a la Ría de Corme e Laxe (mi chica y yo) el 29 de agosto con el objetivo de ver la ardora. Y por supuesto también para recoger muestras y examinarlas en el IEO de Vigo.
Aquel mismo día de mañana observamos una marea roja en la playa de Laxe. O eso parecía…porque al acercarnos resultó ser una acumulación de macroalgas de aspecto plumoso, muy finas. Teñían la orilla de color vino tinto pero no había ninguna proliferación de fitoplancton…
Playa de Laxe (29-VIII-2021). Autora: Mónica Pazos
Una de las playas que citaron en prensa para ver ardora era la de Rebordelo (Cabana de Bergantiños), a 5 minutos en coche de Laxe. Durante la tarde escuchamos conversaciones en terrazas y restaurantes donde hablaban también sobre ella. Así que fuimos a Rebordelo la noche del 29 al 30 de agosto.
A la playa se llega por una carretera estrecha y muy empinada. Mientras discutíamos si habría suerte nos sacó de dudas un fulgor azul a lo lejos. A las 22:30 llegamos andando al arenal y para entonces el acceso ya estaba atascado de coches haciendo cola para aparcar.
La orilla de la playa estaba llena de gente con móviles y cámaras registrando el espectáculo de las olas iluminadas por la ardora.
Pasamos allí casi dos horas. Me bañé con precaución: el mar estaba en calma y el agua fresca pero soportable. El más mínimo movimiento hacía que la ardora iluminase el mar mientras rompían olas azules en la ensenada y en la orilla.
No conseguí grabarla pero dada la cantidad de imágenes y vídeos que se publican lo importante es disfrutar del espéctaculo ¡¡y apagad los flashes por favor!! ¡¡y las linternas hacia el suelo por favor!!.
El desfile de coches iluminaba la playa ocultando la bioluminiscencia y hasta que no paró un poco el tráfico no se pudo apreciar la ardora en todo esplendor.
Cuando regresamos a Laxe revisé las muestras de Rebordelo y a simple vista no veía nada en el agua. No tenían color y tampoco bolitas de Noctilucas. Mantuve las muestras en un lugar fresco y al día siguiente las observé al microscopio en el IEO de Vigo.
Confirmé que no había ni una Noctiluca pero bullían multitud de dinoflagelados, principalmente del género Alexandrium. A bote pronto había dos tipos de células según su tamaño: A. minutum (pequeñito y escaso) y un Alexandrium gordito y redondo que llenaba la muestra y danzaba sin parar.
También había Lingulodinium polyedra de mayor tamaño todavía, poco abundante pero fácil de distinguir por su aspecto acorazado y poliédrico. Muchas especies de Alexandrium (excepto A. minutum) producen bioluminiscencia ¡y Lingulodinium también! Así que todo encajaba.
Faltaba confirmar qué especie de Alexandrium dominaba la muestra. La principal sospechosa era A. tamarense aunque en vivo no hay forma de distinguirla (al menos yo) de otra muy similar: A. ostenfeldii.
Tiñendo sus placas de celulosa pueden discriminarse mediante epifluorescencia gracias a un poro situado entre dos de sus placas. Si dicho poro es diminuto (o no lo puedes ver) se trata de A. tamarense. Y así fue: las células que conseguí observar en la posición adecuada cumplían esa condición.
Células de A. tamarense aisladas de la ardora de Rebordelo. La flecha blanca indica el poro del que les hablé (Vp; no visible en la imagen de la izquierda). Imagen de epifluorescencia obtenida en el IEO de Vigo. Autor: F. Rodríguez
Días después las secuencias genéticas (LSU y ITS rADN) de dos células confirmaron que se trataba de Alexandrium tamarense. Así que dicha especie es seguramente la que produce la ardora en Rebordelo y otras playas cercanas…y me atrevo a decir que en Carnota también.
En la Ría de Corme e Laxe no hay bateas de mejillón. El Instituto Tecnológico para el Control del Medio Marino de Galicia (INTECMAR) posee allí una estación de muestreo (C5) de condiciones ambientales y fitoplancton. Está localizada en Ponteceso, en la desembocadura del río Anllóns en el interior de la Ría. Y a posteriori, en los recuentos de fitoplancton de la semana siguiente destacaba Alexandrium spp. con más de 85.000 células/litro.
Alexandrium tamarense no es tóxico. Aunque si hacen una búsqueda en internet descubrirán que se cita como especie tóxica. La confusión tiene que ver con el hecho de que se denominaron como A. tamarense lo que en realidad eran varias especies de Alexandrium.
A menos que seas un experto en morfología resulta francamente difícil diferenciar sin atisbo de dudas unas cuantas especies de este género. Dado el esfuerzo (y tiempo) que supone identificarlas suelen agruparse bajo la etiqueta de Alexandrium spp., y durante décadas se habló del «complejo de especies A. tamarense» para englobarlas.
En 2014 John y col. publicaron un estudio en el que por fin asignaban nombres válidos a lo que hasta entonces eran Alexandrium tamarense de los grupos I a V. Si quieren ampliar detalles ya conté esta historia en La importancia de llamarse Alexandrium.
¿Cuál es el verdadero Alexandrium tamarense? pues el antiguo grupo III, que corresponde con organismos aislados de la localidad tipo (donde se describió) y que también es el de Galicia.
Esta especie no es tóxica, a diferencia de otras del antiguo «complejo tamarense» (citados como A. tamarense tóxicos en estudios anteriores a 2014).
Marie V. Lebour describió A. tamarense en 1925. Y lo hizo con estas palabras…
(Goniaulax tamarensis) This little species was found up the river Tamar in estuarine water. Cell roundish, rather longer than broad. No apical horn […] Found only in the river Tamar estuary, near Plymouth.
The dinoflagellates of Northern Seas (Lebour, 1925)
Lebour mercería por sí sola una entrada aparte. Fue una ilustre bióloga que estudió diferentes organismos planctónicos, no solo microalgas sino también copépodos y fases larvarias de peces, moluscos y crustáceos.
Comenzó estudiando el microplancton en el Plymouth Marine Laboratory cuando ya tenía 39 años y allí fue donde desarrolló toda su carrera.
Sus libros sobre dinoflagelados y diatomeas fueron los primeros en lengua inglesa. En concreto «The Dinoflagellates of the Northern Seas«, que incluía entre otras la descripción de A. tamarense e ilustraciones propias (no se había descubierto el método de tinción de placas que les mencionaba antes, así que tiene un mérito impresionante con los medios de la época).
Desembocadura del río Anllóns en la Ría de Corme e Laxe. Autor: D. Lema. Fuente: minube
Alexandrium tamarense es una especie estuarina así que encontrarla en la playa de Rebordelo –y con total seguridad en las demás playas de la Ría de Corme e Laxe donde hay ardora– no es ninguna sorpresa.
La combinación del aumento de temperatura en verano y los aportes de agua dulce (menos densa) favorecen la estratificación de la capa superficial con abundantes nutrientes y luz, impulsando el crecimiento de las poblaciones de A. tamarense.
El río Anllóns desemboca en una amplia zona de marisma y en la imagen de satélite que verán a continuación se observan señales intensas de clorofila, a las que deben contribuir las poblaciones de A. tamarense.
Si fuesen Noctilucas no veríamos esa señal porque son heterótrofas (no contienen clorofila propia, aparte de la de sus presas a medio digerir).
Imagen de la desembocadura del río Anllóns, en la Ría de Corme e Laxe (Sentinel 2, tratada con un índice normalizado de clorofila). Autor: Jorge Hernández.
La Ría de Corme e Laxe es casi «de bolsillo» si la comparamos con otras muchas rías gallegas.
Desde el puerto de Laxe la podemos contemplar en buena parte…
Puerto y playa de Laxe. Autor: F. Rodríguez
No es de extrañar que la influencia del Anllóns favorezca el crecimiento de especies estuarinas como A. tamarense en varias zonas de dicha ría, manteniendo poblaciones locales y estimulando su proliferación estival año tras año.
A ello seguramente contribuye también la formación de quistes por esta especie y su permanencia en los sedimentos para «despertar» cuando las condiciones son idóneas.
La ardora en la playa de Carnota también es común durante el verano tal como asegura mi compañera Pilar Rial, asidua del lugar. Y adivinen…a la zona norte de esta inmensa playa llegan aportes de agua dulce que desembocan en la playa «Boca do Río». De hecho, en la colección de cultivos CCVIEO tenemos un A. tamarense aislado de Carnota de una muestra estival recogida por Pilar.
Carnota es la playa más grande de Galicia. Fuente: El Español
El interés que despierta la bioluminiscencia desde el punto de vista turístico ha crecido muchísimo a raíz de las imágenes y vídeos en estos últimos años. Incluso nos llegan consultas al IEO, directamente o a través de la Xunta de Galicia, de personas que quieren visitar Galicia y saber dónde ver la ardora.
No se puede saber con total certeza dónde y cuándo va a haber un mar de ardora espectacular.
Pero sí es posible tener más probabilidades de verlo con información actualizada. Ahora mismo sólo funciona el boca/oreja, las noticias de prensa y los avisos en redes sociales…
Pero veo posible (e interesante por varios motivos) desarrollar una aplicación móvil basada en «ciencia ciudadana» que registre avistamientos de ardora geolocalizados por particulares. Esta aplicación tendría además interés científico: si los datos se recogen de forma continuada a lo largo del tiempo podríamos estudiar tendencias espacio-temporales y relacionarlas con los organismos responsables. Y también obtener datos muy valiosos de cara a desarrollar herramientas predictivas.
Por último y no menos importante: todo esto debería ir acompañado de información adecuada y respeto por los valores medioambientales del entorno.
Volviendo al principio, las manchas anaranjadas de Noctiluca son fáciles de reconocer y dan pistas para buscar playas y zonas de costa con ardora en la/s noche/s siguiente/s. Pero aunque las veas de día, las corrientes puede que las desplacen y te lleves un chasco de noche. Lo mejor es tener paciencia y visitar varios lugares en los alrededores…
En el caso de A Costa da Morte y A. tamarense no tengo noticias deque se vean mareas rojas durante el verano. ¡Ni siquiera yo mismo lo comprobé en esta ocasión! Pero si alguien de ustedes tiene información les agradecería que dejen aquí sus comentarios ¡muchas gracias!
Rebordelo (26-VIII-2021, 4 días antes de la ardora de esta entrada). Autor: Antonio Fuentes Lema
NOTA:¡Pues sí que había marea roja en Rebordelo! el 8 de septiembre de 2021, Antonio Fuentes Lema (@Tonhox4), publicó en twitter varias imágenes de la playa, tomadas el 26 de agosto, con el agua teñida de color marrón.
Los dinoflagelados fotosintéticos con peridinina tiñen el mar de color marrón-rojizo, muy diferente al naranja de las Noctilucas que proliferan en nuestras costas.
Agradecimientos: a todos los que compartisteis imágenes conmigo para elaborar esta entrada, en especial a Jorge por la imagen de Sentinel 2 y a Antonio por confirmar la marea roja en Rebordelo.
Y gracias también a Basti (CACTI, UdV) que siempre hace lo posible por analizar y enviar las secuencias lo antes posible.
Referencias:
Dolan JR. Pioneers of plankton research: Marie Lebour (1876–1971) J. Plankton Res. 1–4. (2021).
John U. y col. Formal revision of the Alexandrium tamarense species complex (Dinophyceae) taxonomy: the introduction of five species with emphasis on molecular-based (rDNA) classification. Protist 165(6):779-804. (2014).
Lebour MV. The Dinoflagellates of the Northern Seas. Plymouth (UK), Marine Biological Association of the United Kingdom, 250pp. (1925).
https://fitopasion.com/wp-content/uploads/2021/09/1630940609756.jpg12001600Francisco Rodriguezhttps://fitopasion.com/wp-content/uploads/2021/06/logo-FITOPASION.pngFrancisco Rodriguez2021-09-06 20:10:402021-09-13 16:43:28No es Noctiluca todo lo que reluce
Conocí Brest durante un congreso en el 2000 y la ciudad me dejó una impresión algo extraña, como si estuviese en una maqueta a tamaño real. Todo muy ordenado y en su sitio.
Pronto supe que Brest había sido arrasada (literalmente) durante los bombardeos en la Segunda Guerra Mundial. Así que aquella sensación que desprendían sus calles emanaba de algo muy real.
Panorámica de Brest desde el castillo medieval. Autor: Getty. Fuente: France Bleu
Con los años descubrí que Bretaña es una región llena de poblaciones con encanto: casas, castillos y edificios singulares que te transportan siglos atrás en el tiempo.
Rennes, Saint Malo y Lorient son ejemplos de otras localidades bretonas destruidas durante la contienda. Pero el caso de Brest fue especial: se trataba de un importante puerto militar ocupado por los nazis en 1940 y donde construyeron incluso una base de submarinos. Allí se hicieron fuertes durante 4 años y no lo dejaron por iniciativa propia…
El río Penfeld (Brest) en octubre 1945. Autora: Gilda Priol. Fuente: brest3945
Brest sufrió bombardeos continuos (¡más de 300!) por parte de los aliados hasta su liberación en 1944. El asedio fue devastador y cuando las tropas estadounidenses alzaron su bandera lo hicieron entre ruinas, en un paisaje postapocalíptico. Solo resistieron buena parte del castillo medieval y el Museo de la Marina en su interior.
Neutralización de una mina alemana en la bahía de Brest (15 septiembre 2020). Autor: Marina Nacional. Fuente: Le Télégramme
La reconstrucción no fue nada fácil.
Hoy en día Brest es una ciudad de aspecto moderno sin casco histórico en la que siguen apareciendo obuses, minas y bombas de aviación (tanto en el subsuelo como en su bahía).
En ella viven unas 140.000 personas y (por si les interesa) está hermanada con A Coruña.
Desde finales del s.XX en la bahía de Brest se ha observado un aumento gradual del dinoflagelado Alexandrium minutum, productor como ustedes saben de toxinas paralizantes (saxitoxinas). Su primer bloom en Bretaña se detectó en 1987, aunque más al sur, en la bahía de Vilaine…
En la bahía de Brest su presencia fue residual hasta 2008. Pero en 2009 explotó un bloom y en 2012 provocó las primeras prohibiciones en la comercialización de marisco (superando 10 veces los niveles permitidos de saxitoxinas). Para entonces sus proliferaciones ya se habían extendido a otros estuarios de la costa atlántica francesa, donde sigue ocasionando cierres frecuentes en el sector acuícola.
Ante una situación así cabe preguntarse si A. minutum es la excepción dentro de un ecosistema estable o si su dominio reciente refleja cambios generales en las comunidades.
Marea roja de Alexandrium minutum en el estuario del Penzé (Bretaña). Fuente: Ifremer.
Entonces…¿ha cambiado el fitoplancton de la bahía de Brest? Pues la respuesta es SÍ.
Ahora vamos con el cuándo, cómo y por qué…
Las series históricas de fitoplancton apenas tienen algunas décadas y no ofrecen amplias perspectivas (excepto p.ej. el CPR survey del Reino Unido, iniciado en 1931).
Pero existen alternativas. La detección de formas de resistencia (quistes) en el fondo marino y los análisis de ADN ambiental permiten averiguar si un dinoflagelado tóxico como A. minutum es nuevo en el barrio o un componente habitual del plancton.
Pues bien. Un primer estudio en 1993 (Erard-Le Denn y col.) no detectó quistes de A. minutum en sedimentos de la bahía de Brest anteriores a 1990. Pero Siano y col. (2021) han dado un paso más allá, muestreando 3 zonas en dicha bahía. Y lo que han hecho es analizar el ADN ambiental en secciones del lecho marino (desde la superficie hasta un máximo de 12 m de profundidad) para reconstruir las paleocomunidades de protistas (eucariotas unicelulares antiguos).
Con esto han conseguido recrear su composición, microalgas incluidas, desde el presente hasta la Edad Media (1121±149). Sólo así es posible conocer el punto de partida en épocas preindustriales y abordar cuestiones tan interesantes como las que verán a continuación…
Durante los siglos que vieron los reinados de Francisco I (1515-1547), el Rey Sol, Luis XIV (1643-1715) o el último Bonaparte (Napoleón III, 1852-1870), el fitoplancton en Brest permaneció impasible tanto al devenir de Francia como al de la humanidad en general.
Por aquel entonces, en la bahía de Brest dominaban dinoflagelados del orden Suessiales (Pelagodinium y Biecheleria/Protodinium). Los Suessiales no producen toxinas y se caracterizan por poseer muchas placas celulares en relación a los demás dinoflagelados. ¿Para qué? Pues vaya usted a saber…
Luego, entre los estramenópilos –que incluyen protistas heterótrofos y diatomeas-, reinaban los primeros.
Los cambios en el fitoplancton de la bahía de Brest llegaron a mediados del s.XX.
Las comunidades de dinoflagelados cambiaron drásticamente durante la década de 1940. Los «históricos» Suessiales fueron sustituidos por un nuevo orden, Gonyaulacales, primero por el género Gonyaulax y a partir de los 80′ Alexandrium y Heterocapsa (orden Peridiniales).
En cuanto a los estramenópilos, el reinado de los organismos heterótrofos dio paso en los 1940’s y 1950’s a las diatomeas entre las cuales destacaba Chaetoceros hasta los 1980’s. Luego, desde los 1990’s, le destronó el género Thalassiosira entre otros.
¿Qué sucedió en esos periodos del s.XX (1940’s y 1980’s) para que el fitoplancton sufriese cambios irreversibles?
Pues una frenética actividad industrial ligada a la ocupación nazi entre 1940-44 y la caída de ¡30.000 toneladas de bombas! sobre la ciudad que seguramente contaminaron la bahía (directamente y a través de aguas continentales).
Así describió aquel infierno el poema «Barbara»: «Sous cette pluie de fer / De feu d’acier de sang» (bajo esta lluvia de hierro / acero fuego de sangre) (J. Prévert).
Resumen gráfico de los resultados de paleocomunidades en Brest. Autor: Siano y col. (2021). Fuente: x-mol.com
Siano y col. confiesan la dificultad de conocer con precisión la composición metálica de los proyectiles pero las anomalías de plomo y cromo en los sedimentos de Brest coinciden con las de Pearl Harbor (EEUU) tras el violento bombardeo de la aviación japonesa.
¿Y los cambios en los 1980’s y 1990’s? En este caso estarían relacionados con el desequilibrio de la proporción nitrógeno/fósforo (N/P) en la bahía de Brest.
El motivo son los nitratos procedentes de fertilizantes y la renovada actividad agrícola a partir de 1950-60’s. Su contínuo desarrollo en la segunda mitad del s.XX llevó a doblar sus niveles en los ríos Aulne y Elorn desde los 70’s a los 90’s.
Las alteraciones antropogénicas disminuyen también la proporción de silicatos (Si) frente a N y P, con otras consecuencias para la composición del fitoplancton: la productividad de diatomeas puede perder peso frente a otros grupos, incluyendo dinoflagelados tóxicos.
A pesar de todo, la bahía de Brest resistió bien a los blooms de dinoflagelados tóxicos hasta la última década. Y para explicarlo existe una hipótesis curiosa: una bomba biológica de silicato relacionada con un organismo invasor.
Verán. Tras la 2ª Guerra Mundial se introdujo el cultivo de ostras del Pacífico (Crassostrea gigas) y con ellas llegó otra especie invasora sin valor económico: el gasterópodo Crepidula fornicata.
Las poblaciones de Crepidula llegaron a ocupar amplias zonas en el fondo de la bahía de Brest…
Hubo planes para erradicarlas y hace 10 años (cuando empezaron a retroceder, no se sabe por qué) eran una grave amenaza para la acuicultura al competir con bivalvos de interés comercial como las vieiras (Pecten maximus).
Pero Crepidulatambién hace otra cosa. Filtran agua como si no hubiese un mañana y producen biodepósitos enriquecidos en silicatos que sedimentan en la bahía un elemento fundamental para las diatomeas.
Así, después del bloom de diatomeas en primavera, los biodepósitos de Crepidula retendrían silicatos que se disolverían luego en el agua, facilitando el crecimiento de diatomeas durante el verano.
A comienzos del s.XXI, resultados experimentales y de modelos apoyaron esta hipótesis «Si/Crepidula», indicando que su erradicación aumentaría la probabilidad de proliferaciones de dinoflagelados tóxicos por limitación de silicatos durante el verano.
En la última década Crepidula ha retrocedido mientras que blooms tóxicos como los de A. minutum se han hecho recurrentes, así que es buen momento para reevaluar dicha hipótesis con las comunidades y balances biogeoquímicos actuales.
El estudio de paleocomunidades en la bahía de Brest (Siano y col.) demuestra que la influencia humana en la bahía de Brest transformó las condiciones ambientales preindustriales alterando el fitoplancton. Y que dichas condiciones también cambian si introducimos «ingenieros del ecosistema» como Crepidula u otras especies invasoras.
Así pues, nada volvió a ser igual después de la guerra. Tanto para las personas como para el fitoplancton…
En 1962 se publicó el álbum «Ives Montand chante Jacques Prévert» que incluía un emocionante poema, «Barbara«, escrito por Prévert en 1946.
Con él comienza y termina esta entrada, para que no olvidemos que la guerra es una idiotez que lo arrebata todo menos el dolor y los recuerdos de quienes sobrevivieron a ella…
Agradecimientos: a Marc Long por enviarme el artículo de Raffaele Siano y col.
Referencias:
Chapelle A. y col. The bay of Brest (France), a new risky site for toxic Alexandrium minutum blooms and PSP shellfish contamination. Harmful algae news 51:4-5 (2015).
Erard-Le Denn E. y col. In: Smayda T.J. & Shimizu Y. (Eds.). Toxic Phytoplankton in the Sea. Elsevier Science Publisher, pp. 109-114 (1993).
Raho N. y col. Biecheleria tirezensis sp. nov. (Dinophyceae, Suessiales), a new halotolerant dinoflagellate species isolated from the athalassohaline Tirez natural pond in Spain. Eur. J. Phycol. 53:99-113 (2018).
Ragueneau O. y col. The Impossible Sustainability of the Bay of Brest? Fifty Years of Ecosystem Changes, Interdisciplinary Knowledge Construction and Key Questions at the Science-Policy-Community Interface. Front. Mar. Sci. 5:124 (2018).
Siano R. y col. Sediment archives reveal irreversible shifts in plankton communities after World War II and agricultural pollution. Curr. Biol. 31:1–8 (2021).
Stiger-Pouvreau, P. & Thouzeau, G. Marine Species Introduced on the French Channel-Atlantic Coasts: A Review of Main Biological Invasions and Impacts. Open Journal of Ecology 5:227-257 (2015).
https://fitopasion.com/wp-content/uploads/2021/05/dl_mlg_imgp0928-modifier05x_bd.jpg9981328Francisco Rodriguezhttps://fitopasion.com/wp-content/uploads/2021/06/logo-FITOPASION.pngFrancisco Rodriguez2021-05-15 12:40:462021-07-10 14:50:50Después de la guerra
Junio de 2002. Una tubería gotea y forma un charco en la acera del campus de la Universidad de Aveiro. En él se agitan dinoflagelados diminutos (Esoptrodinium) a velocidad de vértigo. Luego, cuando el charco se evapora, descansan en el sedimento.
Podrían ser los únicos dinoflagelados «terrestres» que conocemos…
Esoptrodinium gemma. Abreviaturas: Núcleo (N); Cloroplastos (Ch); Mancha ocular (E); Vacuola digestiva (Fv). Los quistes (imagen d) están mezclados con un cultivo de Chlamydomonas. Fuente: Calado y col. (2006).
Pero antes de tratar sobre ellos saltaremos desde Portugal al otro lado de un charco más grande…
Bosque subtropical de Iguazú. Autor: F. Rodríguez
Los bosques lluviosos tropicales de América se denominan Neotropicales y son los mayores del mundo, con una superficie similar a la de Argentina.
Cumplen una función muy importante en la regulación del clima porque aseguran precipitaciones regulares, amortiguan los efectos de las inundaciones, sequías y erosión, etc.
También representan un inmenso depósito de carbono aunque al contrario de la creencia popular su producción neta de oxígeno es despreciable.
La razón es que el oxígeno que liberan durante la fotosíntesis se consume en la fotorrespiración de la propia vegetación y en la descomposición de la materia orgánica.
El O2 que respiramos se lo debemos básicamente al fitoplancton (cianobacterias y microalgas), que comenzó a producirlo cientos de millones de años antes de que hubiesen brotes verdes sobre la tierra.
En los bosques tropicales se calcula que viven el 50-70% de las especies de seres vivos. Pero esa diversidad macroscópica ¿la encontraríamos también a escala microscópica?
Animales varios en los bosques de Iguazú. Ninguno tan interactivo como los coatís (columna izquierda, centro). Autor: F. Rodríguez
Los protistas son seres eucariotasunicelulares entre los que se cuentan las microalgas.Ni animales, ni plantas, los protistas ocupan un reino aparte. Nunca está de más recordarlo…
Pues bien. Mahé y col. (2017) estudiaron la diversidad de protistas en muestras de suelo en bosques Neotropicales de Ecuador, Panamá y Costa Rica (de tipo arenoso, fangoso o secas en lo alto de colinas) y obtuvieron –mediante secuenciación masiva– secuencias de protistas para una región de referencia (V4) en el gen 18S rARN.
Antes de continuar, explicaré un poco de qué va la técnica que usaron...
Los genes ribosomales están presentes en todas las células(razonablemente conservadosdado su rol esencialen el metabolismo) por lo que sirven de marcadores universales para estudiar la biodiversidad y reconstruir las relaciones evolutivas y taxonómicas entre los seres vivos.
Acostumbramos a ver las secuencias de ADN como una serie de bases nucleotídicas (A, T, G, C), pero si representásemos la información útil que contiene el gen 18S rARN para dichos estudios obtendríamos esto en dinoflagelados…
Gráfico de entropía (variabilidad) a lo largo de 1815 bases del gen 18S rARN en 77 especies de dinoflagelados, con las 8 regiones hipervariables. Fuente: Ki (2012).
…y esto otro en copépodos…¿se parecen mucho verdad?
Gráfico de entropía (variabilidad) en el gen 18S rARN en 184 especies de copépodos. Fuente: Wu y col. (2015).
V4 es una región hipervariable del gen 18S rARN, la mayor entre las 8 que existen en dicho gen, con 350-450 bases.
Esa combinación de conservación inherente al 18S rARN y de regiones hipervariables (presentes en todos los organismos) es lo que permite diseñar cebadores universales* para reconstruir las relaciones evolutivas en eucariotas, llegando incluso a nivel de especie.
*Los cebadores son fragmentos cortos de bases nucleotídicas que señalan a la enzima polimerasa el lugar de inicio para amplificar fragmentos de ADN.Universales quiere decir que sirven para que dicha enzima sintetice el ADN de cualquier organismo para la región objetivo (en este caso la región V4 – 18S rARN). En microalgas tanto V4 como V9 ofrecen resultados comparables.
Sigamos. Mahé y col. (2017) emplearon el método Illumina de secuenciación masiva para obtener ¡50,1 millones! de secuencias V4 de protistas. El equipo que usaron permite amplificar con fiabilidad fragmentos de ADN de alrededor de 500 bases. Aquí van sus resultados…
Identificación y abundancia relativa de protistas en el suelo de 3 bosques neotropicales (secuencias totales (reads) y UTOs (en inglés OTUs)). Fuente: Mahé y col. (2017)
Los protistas eran hiperdiversos y dominaban con mucho los parásitos de animales (Apicomplexa), principalmente aquellos de invertebrados. En concreto el 84% de las secuencias y la mitad de UTOs(Unidad Taxonómica Operacional: todo aquello que no quede «sin clasificar» y pueda agruparse en un taxón en los resultados finales).
Lo que más nos interesa aquí es que identificaron como microalgas al 4% de UTOsen el suelo (clorofíceas, haptofíceas, rodofíceas y dinoflagelados). ¿Cómo es posible?
Cianobacterias y clorofíceas pueden vivir sobre sustratos rocosos o simbiontes en líquenes sobre diversas superficies; euglenofíceas, crisofíceas y diatomeas también son habituales en suelo terrestre. Pero en el caso de dinoflagelados (y haptofíceas hasta donde yo sé) no se conocen especies cuyo hábitat incluya el suelo terrestre.
De haberlos cabría esperar que su modo de vida y las comunidades fuesen muy diferentes a las acuáticas. Y no fue así...
Tal fue la sorpresa que en un trabajo posterior se revisaron las secuencias de dinoflagelados del suelo neotropical (Gottschling y col., 2020) y encontraron un poco de todo (representantes de especies fotosintéticas y heterótrofas, marinas y de agua dulce, en los órdenes Gymnodiniales, Peridiniales y Suessiales).
La hipótesis, a falta de documentar la existencia inequívoca de dinoflagelados «terrestres», es que hayan llegado desde el aire llevados por el viento.
Y sólo se conoce un candidato a dinoflagelado «terrestre», que por cierto apareció en el estudio de Gottschling y col. (2020): Esoptrodinium.
El del charco de Aveiro ¡sí!
Se trata de un género extrañísimo del orden Tovelliales descrito en 1997 (Javornický).
Y digo extraño porque poseen un cíngulo incompleto (surco transversal que rodea a toda la célula en la mayoría de dinoflagelados) y un pedúnculo como una trompa por el cual ingieren a sus presas.
Se han encontrado en agua dulce y sedimentos terrestres. La mayoría de especies de Esoptrodinium tienen cloroplastos aunque se alimentan de otras microalgas.
Necesitan luz para crecer en el laboratorio (incluso las especies heterótrofas) y son depredadores generalistas: ingieren presas de múltiples grupos con tal de que posean un tamaño adecuado. Los mejores resultados se han obtenido con criptofíceas (Cryptomonas ovata).
Esoptrodinium es el único candidato a dinoflagelado «terrestre» porque se ha descubierto en charcos en el interior de invernaderos, en prados después de llover (lejos de cuerpos de agua estables), o en una acera de Aveiro (Calado y col. 2006).
Además poseen un superpoder muy útil en tierra: resisten más allá de lo imaginableal calor. Y lo sabemos porque a Fawcett & Parrow (2014) se les estropeó el incubador donde crecían los cultivos y sus quistes sobrevivieron ¡2 días a más de 45°C!
Referencias:
Calado A. y col. Ultrastructure and LSU rDNA-based phylogeny of Esoptrodinium gemma (Dinophyceae), with notes on feeding behavior and the description of the flagellar base area of a planozygote. J. Phycol. 42:434–52 (2006).
Fawcett R.C. & Parrow M.W. Mixotrophy and loss of phototrophy among geographic isolates of freshwater Esoptrodinium/Bernardinium sp. (Dinophyceae). J. Phycol. 50:55–70 (2014).
Gottschling M. y col. The windblown: possible explanations for dinophyte DNA in forest soils. Euk. Microbiol. 0:1-6 (2020).
Javornický P. Bernardinium Chodat (Dinophyceae), an athecate dinoflagellate with reverse, right-handed course of the cingulum and transverse flagellum, and Esoptrodinium genus novum, its mirror-symmetrical pendant. Arch. Hydrobiol. Suppl. 122 (Algol. Stud. 87):29–42 (1997).
Ki J.-S. Hypervariable regions (V1–V9) of the dinoflagellate 18S rRNA using a large dataset for marker considerations. J. Appl. Phycol. 24:1035–1043 (2012).
Mahé F. y col. Parasites dominate hyperdiverse soil protist communities in Neotropical rainforests. Nat. Ecol. Evol. 1:0091 (2017).
Wu S. y col. Taxonomic resolutions based on 18S rRNA genes: a case study of subclass Copepoda. PLoS ONE 10(6): e0131498 (2015).
https://fitopasion.com/wp-content/uploads/2020/12/iguazu.jpg19202560Francisco Rodriguezhttps://fitopasion.com/wp-content/uploads/2021/06/logo-FITOPASION.pngFrancisco Rodriguez2021-01-04 18:15:202021-08-18 21:04:16Lo que el viento se llevó
Imagen de portada: Sinophysis canaliculata. Autoras: M. García-Portela/I. Pazos
En verano de 2008, recién llegado al IEO de Vigo, asistí a la charla de una estudiante de doctorado: Laura Escalera Moura. Ella también acababa de llegar -en su caso de una estancia en la Universidad de Hiroshima.-
La tesis de Laura se centraba en dinoflagelados tóxicos del género Dinophysis pero las muestras del cabo Muroto le reservaban una sorpresa de nombre muy similar: Sinophysis.
En concreto Sinophysis canaliculata, una especie bentónica descubierta en 1999 en islas tropicales del Índico.
Lo de «canaliculata» le viene por un corte en la zona central de la célula que recuerda a la ranura de una hucha.
Aunque en vez de monedas está llena de otras «cosas» como veremos a continuación.
La describieron a partir de células aisladas de algas sobre corales muertos en la isla de La Reunión y explicaron que era «heterotrophic, lacking any chloroplasts»(Quod y col. 1999).
Y el cuento quedó así…
Sinophysis canaliculata. Fuente: Quod y col. (1999).
…hasta que en 2008 Laura y sus colegas japoneses iluminaron las Sinophysis bajo un microscopio de epifluorescencia y en el interior de las huchitas se encendió una pléyade de bolitas naranjas.
Ese tipo de fluorescencia revelaba que S. canaliculata poseía ficoeritrinas, unos pigmentos propios de cianobacterias fotosintéticas.
¿Restos de alimento? ¿endosimbiontes?¿qué diantres de cianobacterias eran?
Para responder a esas preguntas estudiaron las Sinophysis usando microscopía electrónica de transmisión (MET) y genética.
Con la primera técnica vieron las tripas de las Sinophysis mediante secciones celulares. Con la segunda el objetivo era averiguar el nombre y apellidos de las cianobacterias.
Las imágenes de MET demostraron que las cianobacterias estaban libres o rodeadas por una membrana doble –y no en vacuolas digestivas– confirmando así que eran endosimbiontes y no restos de comida.
Imagen MET de Sinophysis canaliculata. Los endosimbiontes (Cy) están señalados con flechas. Fuente: Escalera y col. (2011).
La genética sin embargo no pudo señalar ni el género de las cianobacterias. Nadie en el mundo mundial las había aislado antes, libres o endosimbiontes, para examinarlas con detalle y darles un nombre. Probaron a cultivarlas fuera del huésped, pero sin éxito (Escalera y col. 2011).
Y el cuento continuó así…
…hasta que en 2015, muestreando en charcas costeras de Canarias encontramos unas Sinophysis rosadas entre los dinoflagelados bentónicos de varias islas (entre ellas Lanzarote de donde procede este vídeo).
Por aquel entonces otra estudiante de doctorado en nuestro grupo del IEO de Vigo (María García Portela), se puso manos a la obra con ellas.
Una por una llegamos a juntar 400 Sinophysis para averiguar si eran tóxicas o no (y fue que no: al menos los resultados salieron negativos).
Usando microscopía de epifluorescencia y electrónica de barrido (MEB) confirmó que se trataba de S. canaliculatay que poseía cianobacterias en su interior.
Y los resultados genéticos concluyeron que eran iguales a las de Laura.
Que fuesen iguales en Japón y en Canarias apoya que dichas cianobacterias sean endosimbiontes específicos de S. canaliculata…
…aunque su relación es temporal.
A pesar de que teníamos muchos individuos, estos languidecían a ojos vista y no se dividían: subsistían para morir en pocos meses.
Lo mismo les ocurre a especies fotosintéticas de Dinophysis cuando no tienen a su presa (Mesodinium) para reponer los cloroplastos temporales que se degradan (en este caso de criptofíceas).
Existen dinoflagelados tropicales como Ornithocercus, Histioneis o Citharistes, que también poseen cianobacterias endosimbiontes o ectosimbiontes (en estructuras exteriores).
Curiosamente todos ellos son Dinophysiales como Sinophysis y Dinophysis.
Dichos dinoflagelados, al igual que S. canaliculata, obtienen gracias a sus simbiontes una fuente vital de energía y en varios casos se ha demostrado que además fijan nitrógeno atmosférico, una habilidad muy útil en los desiertos oceánicos…
Estos ejemplos de simbiosis cianobacteria-eucariota son fascinantes para comprender cómo puede evolucionar desde el contacto inicial hasta una integración total en el huésped como cloroplastos.
Una de las hipótesis, fijándonos en esos dinoflagelados (o diatomeas) con cianobacterias externas, es que el proceso podría comenzar así y luego ser integradas en la célula (en vez de la ingestión directa que solemos dar por sentada en dinoflagelados).
En el caso de S. canaliculata todo son incógnitas…
No sabemos si sus endosimbiontes fijan nitrógeno ni cómo llegan al interior de su huésped.
Uno de sus lados (tecas) es plano (como habrán observado en el vídeo) y justo ahí poseen el canal que da nombre a la especie. Quién sabe…¡quizás les sirva para zamparse entre otras presas a las cianobacterias!
En una web llamada AQUASYMBIO encontré una ilustración muy bonita en la que pone que nosotros dijimos(García-Portela y col. 2017) que Synechocystis sp. es el endosimbionte de Sinophysis canaliculata.
Pero no es cierto.Synechocystis no está lejos genéticamente de los endosimbiontes de S. canaliculata, incluso su aspecto es similar…pero son propias de agua dulce.
Nosotros sugeríamos que podrían pertenecer al orden Chroococcales (endosimbiontes en algunas diatomeas y que pueden vivir epilíticas en zonas tropicales).
Así que les escribí al buzón para notificar cambios a la web, comentándoles tanto esto como que los primeros en identificarlas no fuimos nosotros sinoEscalera y col. (2011).
Y el cuento termina así, de momento…
Referencias:
Escalera L. y col. Cyanobacterial endosymbionts in the benthic dinoflagellate Sinophysis canaliculata (Dinophysiales, Dinophyceae). Protist 162:304–14 (2011).
García-Portela M. y col. Morphological and molecular study of the cyanobiont-bearing dinoflagellate Sinophysis canaliculata from the Canary Islands (Eastern Central Atlantic). J. Phycol. 53:446-450 (2017).
Gavelis G.S. & Gile G.H. How did cyanobacteria first embark on the path to becoming plastids?: lessons from protist symbioses. FEMS Microbiol. Lett. 365, fny209 (2018).
Quod J.P. y col. Sinophysis canaliculata sp. nov. (Dinophyceae), a new benthic dinoflagellate from western Indian Ocean islands. Phycologia 38:87–91 (1999).
https://fitopasion.com/wp-content/uploads/2020/10/Scanalicanarias.jpg16411689Francisco Rodriguezhttps://fitopasion.com/wp-content/uploads/2021/06/logo-FITOPASION.pngFrancisco Rodriguez2020-10-31 14:42:262021-08-18 21:05:52Una hucha con sorpresa
Imagen de portada: marea roja en Canido (12 de agosto 2020). Autor: F. Rodríguez.
Este verano en Galicia están surgiendo mareas rojas del dinoflagelado Noctiluca scintillans en muchas zonas de las Rías Baixas, tanto en la de Vigo como en Pontevedra.
Mareas rojas acompañadas de mar de ardora porque Noctiluca es bioluminiscente. Pero una cosa no implica a la otra: el fitoplancton que produce mareas rojas puede pertenecer a distintos grupos y especies. Y sólo algunos géneros del grupo de los dinoflagelados poseen especies bioluminiscentes.
La primera noticia de este fenómeno me llegó desde más al sur: de Matosinhos (Portugal), el 22 de julio.
Marea roja de Noctiluca en Matosinhos (Portugal), 22 de julio 2020. Autora: María García Portela.
Y por si fuera poco, de esta marea roja en Portugal hay un vídeo de mar de ardora (también de María García Portela).
Marea roja de Noctiluca en Silgar (26 de julio 2020. Sanxenxo, Ría de Pontevedra). Autor: J. Filgueira.
Días después se observó una marea roja de Noctiluca en Galicia. Fue el 26 de julio en Silgar (Sanxenxo), y en otras zonas del Morrazo según medios locales (La Voz de Galicia, Faro de Vigo).
Se trata de un fenómeno habitual en verano, a menudo entre julio-septiembre…
…y obedece a un ritmo natural en la sucesión del plancton combinado con condiciones ambientales favorables (temperatura, estabilidad del agua y productividad del plancton).
Marea roja de Noctiluca en Bouzas (Vigo), 29 de agosto 2018. Autor: F. Rodríguez
Son tan habituales que si revisan entradas antiguas del blog encontrarán referencias a sus blooms en las Rías Baixas en 2009, 2014, 2017…y en 2018 (fue pequeñita y no la cité en el blog).
Quizás este 2020 haya sido especialmente Noctilucoso. Lo que sí es seguro es que aparecen todos los años (con más o menos intensidad), en diferentes enclaves de las rías gallegas.
En esas entradas antiguas encontrarán respuestas a muchas cuestiones sobre Noctiluca scintillans. Hoy sólo les recordaré que dicha especie forma parte del 50% de dinoflagelados heterótrofos (la otra mitad son fotosintéticos).
Ciclo de vida de Noctiluca scintillans. Fuente: Fukuda & Endoh (2006).
Y también mencionaré que lo que vemos en la marea roja son trofontes, una fase del ciclo de vida de Noctiluca (la célula de gran tamaño #12 en la parte superior del esquema).
Su flotabilidad provoca que los trofontes se concentren en superficie.
La estabilidad en la columna de agua y su escasa capacidad de movimiento llevan a su acumulación y a la potencial aparición de mareas rojas por acción de las corrientes.
Noctiluca contiene pigmentos (carotenoides) que le confieren ese color anaranjado característico y que permiten a menudo identificar (incluso con imágenes macroscópicas) sus proliferaciones.
Noctiluca scintillans (100 aumentos) aisladas de la marea roja de Canido (12/08/2020). Autor: F. Rodríguez.
El 12 de agosto se observaron manchas en la orilla de varios arenales entre Canido y Samil. Recogí una muestra en Samil después de un aviso de Carlos Vales (coordinador de los Servicios de Salvamento en el Concello de Vigo), y cuando iba de camino al IEO me encontré con esto…
Para confirmar que se trata de Noctiluca necesitamos un microscopio. Aunque por su tamaño (0,5-1 mm) pueden verse a simple vista las bolitas semitransparentes que son sus células.
Después de filtrarlas por gravedad, tal y como muestra el vídeo, las células quedan posadas sobre el filtro. Parecen una tarta…
Noctilucas sobre un filtro de fibra de vidrio de 2,5 cm de diámetro. Autor: F. Rodríguez
En mis charlas de divulgación o en los talleres que hacemos en el IEO suele haber siempre dinoflagelados bioluminiscentes para mostrar la ardora en directo.
En esos casos no usamos Noctilucas (difíciles de mantener durante mucho tiempo en laboratorio), sino dinoflagelados fotosintéticos del género Alexandrium. Pertenecen a la colección de cultivos CCVIEO del IEO de Vigo.
La ardora supone un espectáculo que bien merece la pena el esfuerzo de acercarse a una playa de noche. Pero no siempre es posible desplazarse y además el éxito de un cazador de ardoras es impredecible.
Que haya marea roja de Noctiluca no es sinónimo de mucha ardora: las células pueden dispersarse con las corrientes y luego por la noche apenas veremos tímidos destellos o repentinos fulgores en la rompiente.
Imágenes de la marea roja de Noctiluca en la Ría de Pontevedra. Izquierda: Playa de Lagos (Bueu). Autor: Manuel Garci. Derecha: Playa de Bascuas (Sanxenxo). Autor: Mateo Villaverde.
La imagen más bonita de mar de ardora en Vigo que he visto estos días es de Diego Muñoz. La compartió en su página de facebook donde podrán disfrutar también con imágenes y vídeos de las propias Noctilucas.
Mar de ardora en Canido (13 de agosto 2020). Autor: Diego Muñoz.
Aquí les dejo también el enlace al vídeo que grabó Gerardo Fernández para «A Senda do Moucho» en la playa de Alcabre (Vigo), en esa misma madrugada del 13 de agosto. Al día siguiente lo mostró el Faro de Vigo en un artículo de Sandra Penelas: «Resplandor nocturno en la ría de Vigo».
Convertir el mar de ardora en atractivo turístico de las rías gallegas es tentador pero muy arriesgado dado lo azaroso y variable del fenómeno.
Hay destinos turísticos que sí lo ofrecen, como Puerto Rico, pero sus condiciones naturales del Caribe son privilegiadas. Nada que ver. En Galicia habría que hacer un estudio a lo largo del verano en distintos enclaves de las rías para saber si es posible o no «garantizar» buenas observaciones de ardora.
Turistas pasean en bote por la laguna Grande bioluminiscente de Fajardo en Puerto Rico. Autor: Departamento de Recursos Naturales de Puerto Rico y EFE. Fuente: paginasiete.bo
Personalmente veo casi imposible explotar este recurso natural en Galicia. Pero sí el incluirlo como un complemento de otras actividades, aunque sin garantías de éxito.
Otro problema para que el público general disfrute de la ardora es la contaminación lumínica.
Muchos arenales, y ya no digo si están cerca de núcleos de poblacióncomo Samil o Canido, están iluminados de forma excesiva y sin sentido: farolas, locales comerciales y viviendas envían luz hacia el mar dificultando o impidiendo observar la ardora ¿Para qué?
Aún en playas alejadas, puedes llevarte la sorpresa de un chiringuito de playa que ilumina la playa o viviendas con focos dirigidos al mar. Todo un sinsentido además de un gasto inútil que deberíamos reconsiderar para minimizar dicho impacto.
Mar de ardora en Carnota. Autora: Ana García. Fuente: La Voz de Galicia (15-VIII-2020).
Para aportar algo de luz, se me ocurrió escribir una receta en twitter (@Lilestak) para ver una ardora en casa. Primero debemos entender que las células toleran un rango de temperatura determinado (en nuestro caso 15-25 ºC) y que están adaptadas al ciclo de luz/oscuridad natural, clave para la bioluminiscencia.
Al grano. Si encontramos una marea roja de Noctiluca en la playa, la receta para una ardora casera en 5 pasos sería la siguiente:
Una botella de agua mineral pequeña (500 mL).
Llenar 1/3 de la botella con muestra de la marea roja y luego, fuera de la mancha, completar el resto con agua sin color.
Evitar cambios bruscos de temperatura y mantener la botella en un sitio fresco (nada de nevera ¡que te las cargas!).
A eso de las 21 hrs. poner la botella en un lugar oscuro. Mantenerla así hasta las 23 hrs.
A partir de ese momento, en completa oscuridad, agita suavemente la botella (como si quisieras resuspender azúcar del fondo) y voilà, las Noctilucas deberían iluminarse con la vibración del agua. Puedes ver el efecto varias veces, pero deja reposar un poco la muestra para que recuperen intensidad.
Noctilucas en botella. Son de la marea roja de Canido. Autor: F. Rodríguez
Si no brillan es que no han llegado vivas a la noche…
Los principales motivos son un exceso de temperatura o de concentración. Para saber si están vivas basta comprobar si hay una banda anaranjada en superficie: las células sanas.
El resto, de color más apagado y aspecto amorfo, son células muertas que sedimentan y están degradándose.
La temperatura elevada acelera la degradación de la muestra y si están muy concentradas se morirán antes también. De ahí el consejo de diluirlas.
Con suerte podrían durar vivas en casa 1-2 días. Si tienes fácil acceso al mar es buena idea cambiarles el agua. Basta coger otra botella con agua de mar fresca y añadirle la capa superior anaranjada con las Noctilucas vivas.
Por otro lado, es importante tener en cuenta que la bioluminiscencia obedece a un ritmo interno de las células (circadiano). Necesitarán recibir luz durante el día para mantener ese ritmo interno y generar bioluminiscencia por la noche.
Noctiluca scintillans. Autor: Diego Muñoz.
Y esta no aparece de golpe al 100%de intensidad, sino que aumenta gradualmente para decaer luego al llegar el nuevo día.
Así que al inicio de la noche no brillarán mucho aún ¡sólo es cuestión de esperar un poco más!
Agradecimientos: A todos a quienes cito en esta entrada y que han compartido sus imágenes y vídeos para ilustrarla. Muchas gracias a tod@s.
Referencias:
Fukuda Y. & Endoh H. New details from the complete life cycle of the red-tide dinoflagellate Noctiluca scintillans (Ehrenberg) McCartney. Eur. J. Protistol. 42(3):209-19 (2006).
https://fitopasion.com/wp-content/uploads/2020/08/Toralla.jpg5611024Francisco Rodriguezhttps://fitopasion.com/wp-content/uploads/2021/06/logo-FITOPASION.pngFrancisco Rodriguez2020-08-16 17:36:462021-07-14 21:18:13Las Noctilucas son para el verano
Imagen de portada: Sabrina (Billy Wilder, 1954). Fuente: toldbydesign
Después de las 2 entradas de Suso Gago sobre macro- y microplásticos (MP), hoy cerramos la trilogía plástica explicando la interacción entre el fitoplancton y esos materiales que revolucionaron el siglo XX.
En su honor, esta entrada va a ser un poquito vintage.
Empezando por la portada, que escogí como ejemplo de dicha revolución tecnológica. En Sabrina (1954) un empresario de éxito (Humphrey Bogart) se empeña en demostrar divertidamente las propiedades de un plástico ideal (flexible, a prueba de balas e ignífugo)…lo más de lo más...
(M Schilthuizen, 2019). Fuente: Turner.
Décadas después (remake de Sabrina incluido), el impacto de nuestra civilización en el planeta ha dejado una impronta de basura plástica en todos los océanos, incluso en la fosa de las Marianas.
Si esto les aflige, para levantar el ánimo les recomiendo «Darwin viene a la ciudad«. A mí al menos me ha servido para considerar desde una perspectiva más amplia nuestro impacto sobre el planeta.
Su autor explica la evolución de los seres vivos en zonas urbanas, a las que destaca como un elemento más de la naturaleza.
Las especies animales y vegetales del antiguo entorno salvaje (junto a las invasoras), pelean por los recursos y desarrollan adaptaciones para sobrevivir en la jungla urbana.
El ecosistema de las ciudades es dinámico y fragmentado. Ofrece nuevos recursos, estructuras y sustancias –a menudo tóxicas– que afectan a los procesos evolutivos (p.ej. selección natural y sexual), a expensas de la biodiversidad.
Schilthuizen se centra en organismos terrestres, sobre todo en aves. Pero también menciona a los ecosistemas acuáticos y creo que su discurso es aplicable igualmente a plásticos y microalgas.
Así que digannos, por 25 pesetas, algunos ejemplos de interacciones entre plásticos y microalgas. Por ejemplo: la dispersión. 1, 2, 3, responda otra vez.
¡ La dispersión !
Microorganismos sobre plásticos. (A,B,C) A. taylori. (D,E) Quistes desconocidos. (F) Ostreopsis. (G) Prorocentrum. (H,I) diatomeas (J) hidrozoo (K) macroalgas. Fuente: Massó y col. (2003).
Las microalgas bentónicas habitan sobre sustratos inertes (rocas) o vivos (macroalgas): son un miembro más de la biopelícula microbiana.
Al fitoplancton tanto le da colonizar restos de madera, conchas de moluscos, el lomo de una ballena o cachitos de plásticos que flotan a la deriva.
¡Mientras haya luz y nutrientes todo va bien!
A estas microalgas que habitan en las fibras de poliester o tapones de botellas podríamos llamarlas epiplásticas.
Se calcula que en total, sobre los cachitos de plástico en el mar, podrían vivir de 1.000 a 15.000 toneladas de microorganismos.
La basura marina, plásticos incluidos, es un nuevo hábitat para el crecimiento y dispersión del fitoplancton (incluyendo especies nocivas).
Ya lo dijeron Massó y col. (2003) tras estudiar plásticos flotantes, en la costa mediterránea de Cataluña, colonizados por células vegetativas y quistes de dinoflagelados (Ostreopsis, Coolia, Alexandrium taylori…).
Las comunidades epiplásticas son distintas a las del agua que les rodea, aunque en ello influyen varios factores como su antigüedad, la degradación del material o si han pasado…¡por el tracto digestivo de algún animal!
Fuente: Mincer y col. (2016).
A menudo, cuando el plástico llega al mar, las primeras microalgas que colonizan su superficie son diatomeas.
En las imágenes de la derecha podemos ver diatomeas pennadas pegadas a poliestireno expandido (el corcho blanco de los envases) sumergido durante 1 semana en el Atlántico Norte.
No obstante, además de diatomeas, los estudios genéticos revelan que sobre el plástico pueden habitar numerosos grupos: criptofíceas, cocolitofóridos, prasinofíceas, pelagofíceas, etc.
¡ La sedimentación !
Aunque a los que vivimos en la costa pueda parecernos que esto de la basura marina va cada vez a más, los estudios disponibles no confirman una tendencia clara y de hecho su presencia en el océano es muy inferior a la que cabría esperar.
¿Dónde está ese plástico que falta? La respuesta parece ser en el fondo del mar.
En 2014 un estudio de Woodall y col. descubrió que la abundancia de MP en 12 muestreos en el Mediterráneo, Atlántico e Índico era 4 órdenes de magnitud superior a la de la superficie en los giros oceánicos (que como bien saben concentran la basura marina por acción de las corrientes).
Pues bien. El fitoplancton podría jugar un papel importante en ese transporte profundo al adherirse los MP sobre agregados de partículas como los que producen las diatomeas.
Dicho grupo, clave en la producción primaria marina, se caracteriza por producir grandes cantidades de partículas exopoliméricas transparentes (en inglés, TEP). las TEP son pegajosas y agregan a su vez a otras partículas, contribuyendo a la exportación de materia orgánica, CO2 y por qué no, MP, hacia aguas profundas.
Apenas existen trabajos sobre ello, pero Long y col. (2015) demostraron que los agregados celulares de diatomeas y criptofitas incorporan microplásticos aumentando enormemente su velocidad de sedimentación.
Aunque el efecto varía según la naturaleza tanto de los MP (p.ej. densidad y carga superficial) como de los agregados. En el caso de las diatomeas (Chaetoceros neogracile) observaron que los agregados con MP sedimentaban a menor velocidad que los agregados «limpios». Es decir: los plásticos hacían flotar más a las diatomeas.
Esto plantea cuestiones sobre las consecuencias para el plancton y otros organismos que se alimentan de dichos agregados (que forman la llamada nieve marina) así como la eficiencia de la bomba biológica.
Ya ven. Queda mucho por estudiar. Y esto nos da pie a la última respuesta de nuestros concursantes del 1, 2, 3…
¡ La toxicidad !
De entrada hay que decir que la concentración actual de MP en el mar es poco probable que tenga efectos sobre el metabolismo del fitoplancton.
Pero la preocupación sobre este asunto alienta en la actualidad múltiples investigaciones y proyectos para conocer los posibles efectos fisiológicos en microalgas expuestas a concentraciones elevadas de MPy de nanoplásticos (NP: <100 nm).
¿Y qué concluyen esos trabajos? pues que las consecuencias suelen ser neutrales o negativas: liberación de sustancias tóxicas, descenso en el contenido de clorofila, de la actividad fotosintética y del crecimiento; estrés oxidativo, alteraciones metabólicas y morfológicas.
Resumen de efectos de los microplásticos sobre el fitoplancton. Fuente: Correia-Prata y col. (2019).
El tipo de MP y su tamaño son factores importantes a la hora de su interacción con el fitoplancton. En estudios comparativos, los MP con carga positiva y de menor tamaño son más tóxicos para las microalgas.
Sin embargo, los factores responsables de la toxicidad de los MP y de la sensibilidad de determinadas especies o grupos del fitoplancton no están nada claros.
Pero no todo va a ser toxicidad. Los plásticos son polímeros cuyo elemento principal es el carbono y se estima que sus residuos en el mar liberan anualmente 23.000 toneladas de carbono orgánico disuelto.
Esto supone una fuente de energía extra para las comunidades microbianas heterótrofas, con un impacto que sí podría ser significativo –y cada vez mayor– sobre la actividad y la composición de dichos microorganismos y otros miembros del plancton, así como para el ciclo del carbono en el océano (Romera-Castillo y col. 2018).
¡Campaaaaaana y se acabó!
La investigación de los efectos de los MP y NP sobre el fitoplancton es un asunto reciente y de candente actualidad…
…y dada la importancia crucial de los productores primarios –y del plancton en su conjunto para la salud de los ecosistemas- cabe esperar un auténtico boom de proyectos en el futuro que profundicen sobre este tema.
Referencias:
Correia-Prata y col. Effects of microplastics on microalgae populations: A critical review. Sci. Total Environ. 665:400-405 (2019).
González-Fernández C. y col. Do transparent exopolymeric particles (TEP) affect the toxicity of nanoplastics on Chaetoceros neogracile?. Environ. Pollut. 250:873-882 (2019).
Long M. y col. Interactions between microplastics and phytoplankton aggregates: Impact on their respective fates. Mar. Chem. 175:39-46 (2015).
Massó M. y col. Drifting plastic debris as a potential vector for dispersing Harmful Algal Bloom (HAB) species. Sci. Mar. 67:107-111 (2003).
Ortega-Retuerta E. y col. Horizontal and Vertical Distributions of Transparent Exopolymer Particles (TEP) in the NW Mediterranean Sea are Linked to Chlorophyll a and O2 variability. Front. Microbiol. 7:2159 (2017).
Romera-Castillo C. y col. Dissolved organic carbon leaching from plastics stimulates microbial activity in the ocean. Nat. Comm. 9:1430 (2018).
Seoane M. y col. Polystyrene microbeads modulate the energy metabolism of the marine diatom Chaetoceros neogracile. Environ. Pollut. 251:363-371 (2019).
Woodall L.C. y col. The deep sea is a major sink for microplastic debris. R. Soc. Open Sci. 1:140317 (2014).
Yokota K. y col. Finding the missing piece of the aquatic plastic pollution puzzle: Interaction between primary producers and microplastics. Limnol. Oceanogr. Lett. 2:91-104 (2017).
https://fitopasion.com/wp-content/uploads/2020/05/sabrina.jpg392512Francisco Rodriguezhttps://fitopasion.com/wp-content/uploads/2021/06/logo-FITOPASION.pngFrancisco Rodriguez2020-05-22 18:04:312020-05-22 18:04:32Cachitos de plástico y fitoplancton
El 8 de abril de 1940 los ejércitos de Hitler lanzaban una ofensiva sobre la costa noruega, atacando primero Oslo y Trondheim e invadiendo luego todo el país. Fue el comienzo de 5 años de ocupación nazi…
Aquella situación la sufrió en primera persona el biólogo marino Trygve Braarud, dedicado al estudio del fitoplancton en la Universidad de Oslo. Braarud trabajaba en aquel momento con el género de dinoflagelados Peridinium, con especies como P. trochoideum (hoy Scrippsiella trochoidea).
Enrique Balech le pidió muestras para su estudio pero Braarud no pudo enviárselas por culpa de aquellas circunstancias (algo que sí pudieron atender otros colegas como Guillard y Sousa e Silva). Décadas después, en 1974, Balech redescribió el género Protoperidinium para incluir las especies marinas mezcladas antes en Peridinium junto a organismos de agua dulce.
El género Protoperidinium lo creó R.S. Bergh en 1881 y Balech lo redefinió como exclusivamente marino en base a criterios morfológicos comunes (diseño y número de las placas de celulosa) que no observaba en las especies de aguas continentales.
Protoperidinium no realiza fotosíntesis, se alimenta de otros microorganismos por lo que podemos considerarlo un género de animalillos microscópicos.
Los dinoflagelados heterótrofos pueden comer muchas cosas, desde bacterias, microalgas (incluyendo dinoflagelados) a huevos, larvas de zooplancton e incluso los pellets fecales de éste. Capturan el alimento usando 3 estrategias básicas: 1) ingestión directa, 2) usando un pedúnculo o 3) mediante una estructura retráctil a modo de «saco»: el palio.
Esa estrategia es la que utiliza Protoperidinium y es la que le permite alimentarse de presas mucho mayores que él, incluso hasta 10 veces más grandes !!
Los dinoflagelados que ingieren directamente a sus presas lo suelen hacer por zonas concretas, por ejemplo el sulco, una «rendija vertical» en la parte ventral de la célula.
Dinoflagelados con distintos tamaños de sulco: una posible adaptación para ingerir presas de mayor tamaño. Fuente: Jeong y col (2010)
Obviamente eso limita el tamaño máximo de lo que comen. Aunque hay excepciones: Fragilidiumse deforma y separa sus placas para abarcar presas mayores que él.
Otros dinoflagelados como Prorocentrum parecen tener distintas «bocas» y consiguen alimentarse de varios organismos a la vez por medio de diferentes suturas en la célula.
Pero la regla general es que sólo aquellos equipados de un pedúnculo o el palio pueden alimentarse de seres de mayor tamaño.
Y pueden atacar en grupo…
Hace varios años publiqué una entrada con vídeos de Protoperidinium alimentándose de otro dinoflagelado (Tripos fusus), y 3 de ellos arrastrando juntos lo que parecía un huevo de zooplancton. Una lectora llamada Laura me preguntó hace unos días por este asunto y su comentario motivó que escribiese esta entrada, así que muchas gracias por seguir el blog y participar con vuestros comentarios !
Aquí van tres nuevos vídeos de Protoperidinium, pertenecientes a aquellas muestras de otoño de 2013, donde le podemos observar solito y con presas (dinoflagelados en este caso)…
Cuando localiza una posible presa, Protoperidinium la merodea con una especie de danza en espiral que indica que está preparándose para desplegar el palio. Pero justo antes de eso «pesca» a su presa extendiendo un filamento en el extremo de un filopodio (una delgada proyección de su citoplasma).
Protoperidinium spinosum alimentándose de una diatomea tubular. (MB) es la cesta microtubular que comunica con el palio. Fuente: Jacobson & Anderson (1992).
El palio es un tipo de pseudópodo (lamelipodio) que consiste en una prolongación laminar del citoplasma dotada de una intrincada red de canales y vesículas.
En la zona media de la célula de Protoperidinium existe una estructura ovalada («cesta microtubular», según Jacobson & Anderson 1992), desde la que se proyecta el palio, y cuyo contenido citoplasmático (o «palioplasma» mejor dicho) es continuo entre ambos.
¿Por dónde sale al exterior el palio? pues por la zona del sulco aunque éste, como el resto de la superficie de Protoperidinium, está tapizado de placas.
Pero una de ellas, la placa sulcal media, hialina y muy delgada, es la candidata a permitir las idas y venidas del palio.
Sobre dicha placa el propio Balech se aventuraba en 1974 a comentar que podría desconectarse de las demás para permitir la salida del pseudópodo (el palio).
Él mismo añadía que sólo conocía dicha forma de alimentación en Peridinium (citando el divertido nombre de Peridinium gargantua) pero que hasta la fecha nunca la había observado en Protoperidinium.
Protoperidinium depressum (Dn) alimentándose de una cadena de diatomeas (Dt) usando el palio, en la fase inicial (A) y avanzada donde extiende el palio a lo largo de su presa. Autor: D. M. Jacobson (1987). Fuente: American Journal of Botany
La mayoría de las presas de Protoperidinium suelen ser diatomeas (aunque también, como hemos visto, puede alimentarse de otros dinoflagelados), a las que digiere extracelularmente tras envolverlas en el palio como quien empaqueta su bocadillo en plástico transparente !!
Los contenidos celulares de la presa son digeridos mediante enzimas, succionados y transportados hasta la cesta microtubular en el centro de la célula.
Cuando todo finaliza el palio se retrae liberando los restos sin digerir, como las frústulas silíceas de las diatomeas. El proceso entero de alimentación puede durar entre 5 y 30 minutos.
Y los hay con digestión más lenta. Géneros cercanos a Protoperidinium como Oblea se han llegado a observar dale que te pego hasta 2 horas !!
Referencias:
-Balech E. El género «Protoperidinium» Bergh, 1881 («Peridinium» Ehrenberg, 1831, partim). Revista del Museo Argentino de Ciencias Naturales «Bernardino Rivadavia» Instituto Nacionale de Investigacion de Ciencias Naturales, Hidrobiologia 4(1): 1-79 (1974)
-Jacobson DM & Anderson DM. Ultrastructure of the feeding apparatus and myonemal systema of the heterotrohic dinoflagellate Protoperidinium spinulosum. J. Phycol. 28: 69–82. (1992)
-Jeong HJ, Yoo YD, Kim JS, y col. Growth, feeding and ecological roles of the mixotrophic and heterotrophic dinoflagellates in marine planktonic food webs. Ocean Sci. J. 45:65-91 (2010)
https://fitopasion.com/wp-content/uploads/2017/05/oogie-boogie.jpg283375Francisco Rodriguezhttps://fitopasion.com/wp-content/uploads/2021/06/logo-FITOPASION.pngFrancisco Rodriguez2017-05-28 21:08:452017-05-28 21:09:28El dinoflagelado del saco
Utilizamos cookies propias y de terceros para garantizar el funcionamiento de la web, medir su uso y mejorar nuestros servicios. Puede aceptar todas las cookies, rechazar las no necesarias o configurar sus preferencias. Política de cookies
Configurar cookies
Necesarias para que la web funcione correctamente. No se pueden desactivar desde este panel.
Ayudan a medir el uso del sitio y mejorar sus contenidos.
Permiten publicidad, medición de campañas o personalización de anuncios.
Guardan preferencias o permiten contenido externo no necesario.






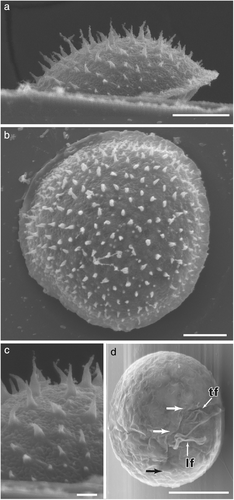

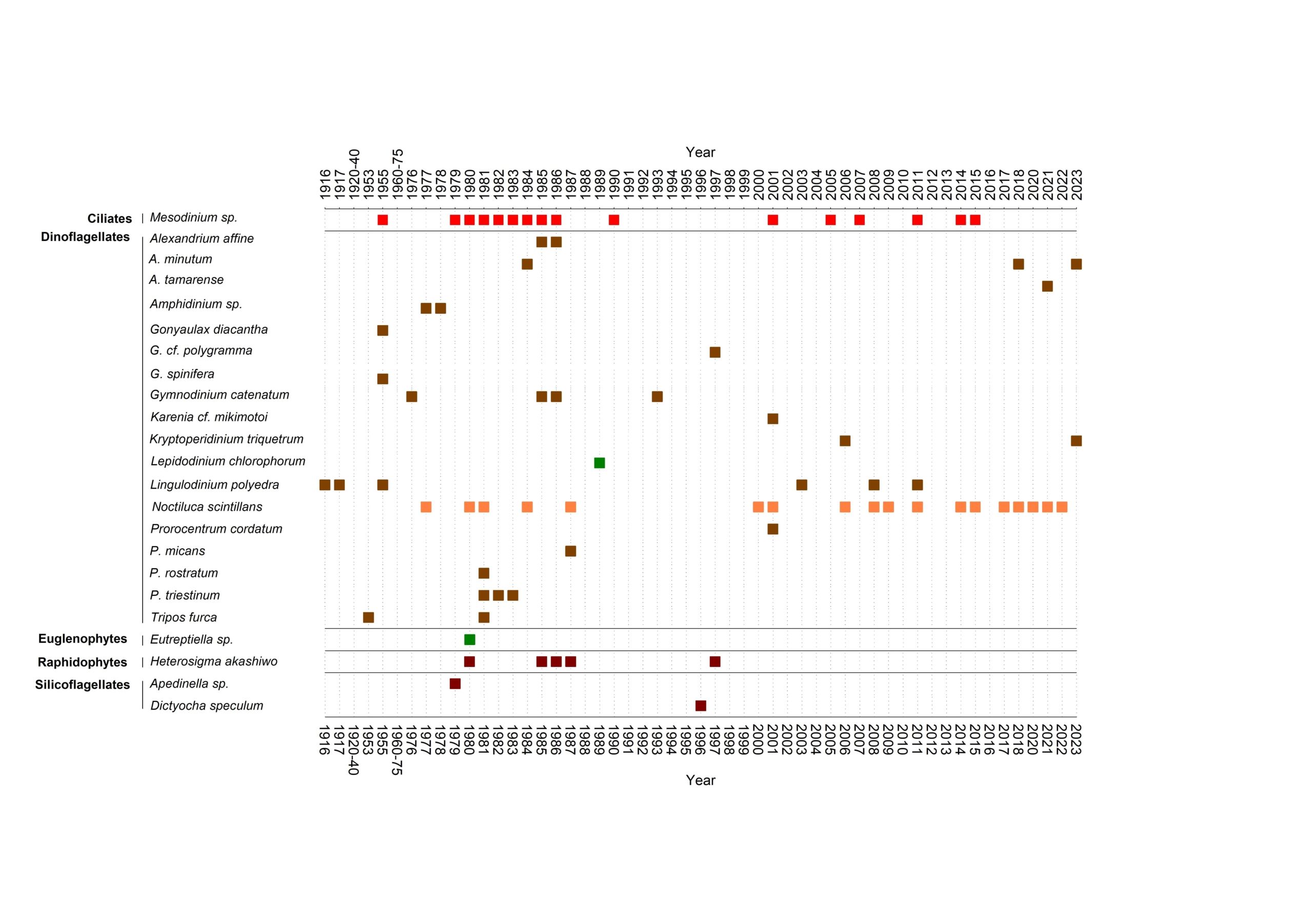





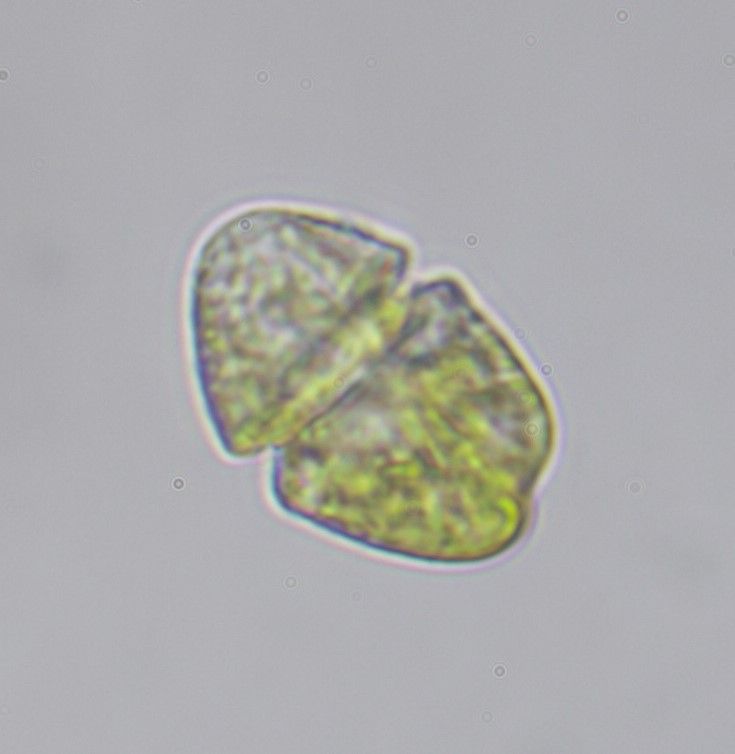















































































{kind=link}