
El nitroplasto
Imagen de portada: Braarudosphaera bigelowi. Autores: Monteiro FM y col. Fuente: Wikimedia Commons
Las tecnologías y objetos de la antigüedad son difíciles o imposibles de interpretar sin imágenes ni documentos. ¿Cómo funcionaban? ¿Para qué servían?
Por ejemplo, en la última entrega de Indiana Jones toda la trama gira alrededor de encontrar el «Dial del Destino». Este artilugio, que en la película permite (ATENCIÓN SPOILER, viajar en el tiempo), se basa en el Mecanismo de Anticitera, un artefacto auténtico encontrado en un pecio y que se cree que servía para predecir posiciones astronómicas. Pero existen objetos de los que no sabemos nada como los dodecaedros romanos.

Dodecaedro romano encontrado en 1939 en Tongeren (Bélgica). Fuente: Wikimedia Commons.
Los dodecaedros romanos poseen tamaños parecidos a pelotas de golf. Suelen ser metálicos y huecos, con agujeros en la superficie de distintos diámetros y protuberancias redondas en los vértices. No se han encontrado testimonios de ningún tipo sobre ellos así que desconocemos para qué servían. En todo caso no parecen herramientas sino algo valioso.
Descubrir nuevas especies de fitoplancton puede ser tan emocionante como toparse con un dodecaedro romano. A primera vista te maravilla su diseño y luego te preguntas ¿Pero esto qué es?!!! Algo así debieron pensar quienes observaron por primera vez una microalga con forma de dodecaedro que casualmente también escondía un misterio que ya imaginaréis por el título de esta entrada.

Fuente: ecartelera
Hablamos de una especie viva así que el misterio y las pistas para llegar a él estaban intactas en su interior. Solo hubo que esperar a desarrollar las tecnologías y conocimientos necesarios.
Para que entendamos la historia primero os presentaré a los personajes: el dodecaedro, un flagelado y una cianobacteria.
-EL DODECAEDRO-
En 1931 una comisión pesquera internacional invita a dos noruegos, Gran y Braarud, a estudiar la oceanografía y el fitoplancton en la bahía de Fundy y el golfo de Maine (EEUU). El objetivo era conocer los efectos que tendría sobre la productividad primaria la construcción de un embalse. Pues bien, en medio de un farragoso trabajo de 150 páginas incluyeron el descubrimiento (por parte de Braarud) de un nuevo cocolitofórido: Pontosphaera bigelowi. Sus células eran dodecaedros cubiertos por 12 cocolitos calcáreos pentagonales (llamarles cocolitos no es correcto ya que por definición son redondos. Lo correcto sería «pentalitos»).
La descripción de aquella especie la despacharon en pocas líneas y con un sencillo esquema. Voici…
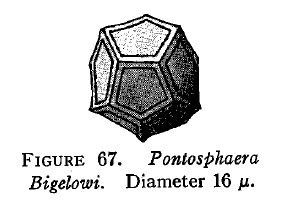
Fuente: Gran & Braarud (1935).
En 1947 otro investigador, Deflandre, vuelve a encontrar los dodecaedros calcáreos en muestras del Atlántico NE (a 645 m de profundidad y 100 km al oeste de Casablanca). Las examinó en detalle y creó una familia taxonómica para estos cocolitofóridos. En homenaje a Braarud renombró Pontosphaera bigelowii como Braarudosphaera bigelowii. Y en su ilustración recoge nuevos elementos como que los pentalitos están compuestos por 5 placas pentagonales.

Fuente: Deflandre (1947).
¿Cómo os quedáis si os digo que Braarudosphaera es un «fósil viviente»?
Los seres vivos son las versiones más exitosas, las mejor adaptadas para transmitir su legado a las siguientes generaciones. Somos el resultado de la evolución y del azar de los tiempos porque un meteorito o una glaciación lo cambian todo y llevan a la extinción al más poderoso de los dinosaurios y por poco a nuestra propia especie, respectivamente.
Pues bien, las especies conocidas como «fósiles vivientes» (término acuñado por Darwin) no son versiones estáticas ajenas a la evolución: NADIE LO ES. Si pudiéramos analizar y comparar su ADN actual con el de sus parientes fósiles encontraríamos más cambios de los que sugiere su aspecto. El secreto de su «longevidad» parece estar, entre otras cosas, en tasas más lentas de evolución frente a otras especies.
Si os pregunto por «fósiles vivientes» pensaréis en celacantos, cangrejos herradura, ginkgos…la realidad (¡¡y esto no lo vereis en las noticias!!) es que las especies de fitoplancton también se extinguen, como todo hijo de vecino, pero el concepto de «fósiles vivientes» también es aplicable a ellas.

Cangrejo herradura del Atlántico (Limulus polyphemus). Autor: Kaldari. Fuente: Wikimedia Commons.
Las microalgas calcáreas como los cocolitofóridos fueron las reinas del mambo durante el Cretácico, periodo geológico que por algo les debe su nombre (hace 145-66 millones de años). En aquel periodo Braarudosphaera formaba blooms en el golfo de México y el mar de Tetis como demuestra su presencia en el registro fósil. Pero luego se estampó el meteorito que arrasó con los dinosaurios y el mambo se terminó.Se extinguieron el 75% de especies marinas y más del 90% de las microalgas calcáreas.
Si os pica la curiosidad por saber cómo eran la Tierra y sus océanos en aquella época seguid este enlace (ya vereis que guay…https://dinosaurpictures.org/ancient-earth#66)
¿Adivinais quién sobrevivió al cataclismo? por supuesto, Braarudosphaera, aunque ya no hace blooms y solo llegaron a nuestros días 2 especies: B. bigelowii y B. magnei (y de esta última casi no sabemos nada).
-EL FLAGELADO-
Viajemos ahora a 1972. Ese año se describe Chrysochromulina parkeae, una nueva haptofita encontrada en aguas al sur de Inglaterra y de Noruega. Se trata de un flagelado desnudo igual que el resto de especies en dicho género. Ni fú ni fá…una cosa sosa que apenas fabrica minúsculas escamas calcáreas.
Pero os recuerdo (guiño, guiño) que los cocolitofóridos como Braarudosphaera también son haptofitas…

Chrysochromulina parkeae. Swedish Meteorological and Hydrological Institute. Copyright © 1996-2006, Mats Kuylenstierna & Bengt Karlson. Fuente: SMHI
-LA CIANOBACTERIA-
Entramos en el siglo XXI y asistimos al BOOM de las técnicas moleculares (una revolución equivalente al reggaeton en la música). Y casi un siglo después gracias a la biología molecular -combinada con otras técnicas- quedaron al descubierto los secretos de Braarudosphaera bigelowii.
Si conoces la función de un gen (o grupo de genes) y este se expresa en muestras naturales puedes confirmar la presencia de dicha función aunque no veas qué organismos la realizan. Así fue como demostraron en 2001 (mediante la expresión de genes nifH relacionados con fijación de nitrógeno), la existencia de cianobacterias marinas unicelulares fijadoras de nitrógeno atmosférico (N2). Esto fue tan novedoso que les valió (y aún les vale) a sus descubridores de la Universidad de California (Zehr y col.) varias publicaciones en Nature y Science.
Hasta entonces se creía que solo cianobacterias filamentosas como Trichodesmium y otras endosimbiontes en diatomeas (p.ej. del género Rhizosolenia y Hemiaulus) eran capaces de absorber N2 y transformarlo en amonio como fuente de nitrógeno. Esto viene a ser una especie de superpoder ya que permite acceder a otra fuente de nutrientes para sobrevivir en aguas oligotróficas.

La diatomea Hemiaulus hauckii. La flecha señala el heterocisto (fijador de nitrógeno) de la cianobacteria endosimbionte (Richelia intracellularis) en color amarillo-naranja mediante epifluorescencia. Fuente: Schouten y col. (2013).
Y ahora se unían a este selecto club unas cianobacterias unicelulares desconocidas (con distribución global) y detectadas en una fracción menor a 10 micras.
En 2008 los mismos investigadores aislaron y concentraron mediante citometría aquellas cianobacterias para estudiar su genoma. Les llamaron «UCYN-A» aunque en otro estudio propusieron llamarlas «Candidatus Atelocyanobacterium thalassa» mientras nadie las describiera formalmente. Primera vez que leo esto de «Candidatus…».
Al grano. Comprobaron que costaba separar a «Candidatus…» de otros microorganismos fotosintéticos y heterótrofos. Al final concentraron una población celular enriquecida en «Candidatus…» y descubrieron que su genoma era muy simple. Este proceso de reducción genética (y metabólica) es el que han sufrido también orgánulos celulares como las mitocondrias y cloroplastos que proceden de antiguos procariotas de vida libre. De hecho sorprendía que pudiera sobrevivir en forma libre con un genoma sin genes esenciales para la fotosíntesis y la fijación de carbono.
Ya tenemos a todos los personajes y ahora es donde convergen en el desenlace final…
Chrysochromulina parkeae es una fase del ciclo de vida de B. bigelowii. Su aspecto no tiene nada que ver pero un estudio genético en 2013 demostró que son practicamente idénticas. Esto es habitual en cocolitofóridos donde suelen alternar 2 fases (morfotipos) que pueden incluir un flagelado móvil (no calcificado: C. parkeae) y una fase inmóvil (calcificada: B. bigelowii).

Braarudosphaera bigelowi. Imágenes de SEM (A-C) y microscopía óptica (D-F). Fuente: Hagino y col. (2013).
Braarudosphaera bigelowii tampoco lo puso fácil porque además de no ser abundante no se pudo cultivar en laboratorio hasta 2021. Pero ya en 2020 hay un estudio (Cabello y col.) donde citan observaciones personales del japonés Hagino (el primero en cultivarla) que confirman esa alternancia de «flagelado» y «dodecaedro».
En 2012 otro estudio de citogenética (citometría + genética; esta vez en Science), identificó a las microalgas que daban positivo en «Candidatus…». La técnica funciona como un control de carretera donde registran el maletero de los coches a medida que pasan (pero usando células y fluorescencia en vez de coches y linternas). Y así fue como descubrieron que aquellas cianobacterias no vivían libres sino en simbiosis con B. bigelowii. Esto resolvía la incógnita de cómo sobrevivir con un genoma tan simple.
Los autores de aquel trabajo sugirieron que «Candidatus…» debía encontrarse fuera de B. bigelowii porque cuando procesaban las muestras se separaban fácilmente. Aunque no las veían y aquella suposición podía estar equivocada. Y así fue porque otros las observaron y descubrieron que estaban dentro.
Al año siguiente, un estudio con microscopía electrónica y genética demostró que en el interior de B. bigelowii había una estructura esférica cuya secuencia genética coincidía con «Candidatus…». Así que estaban dentro como endosimbiontes temporales o incluso un orgánulo celular permanente. Esto último sería un «notición» porque ninguna microalga (que son eucariotas os recuerdo) puede fijar nitrógeno por si misma. Se aprovechan de cianobacterias simbiontes y nadie ha confirmado que sean permanentes.

Braarudosphaera bigelowi (imagen TEM). La estructura esférica señalada con «S» es la cianobacteria «UCYN-A» o «Candidatus…». Fuente: Hagino y col. (2013).
¿Y cuál es la frontera entre un endosimbionte temporal y un orgánulo celular permanente?
Es obligado empezar por Lynn Margulis (de soltera L. Petra Alexander) quien planteó en su histórico trabajo de 1967 «On the Origin of Mitosing Cells» la teoría de que tanto mitocondrias como cloroplastos eran orgánulos celulares permanentes derivados de endosimbiontes.
En el caso que nos ocupa, cianobacterias endosimbiontes, un trabajo de 2009 planteó la hipótesis «del equipaje» para sugerir que cuanto más difícil sea encontrar al simbionte más probable será que su asociación evolucione desde temporal a orgánulo permanente. Nadie devolvería una pepita de oro a la arena de la playa y siguiendo esta hipótesis B. bigelowii preferirá cargar con «Candidatus…» como equipaje (transmisión vertical) para que su descendencia no pierda el superpoder de fijar nitrógeno atmosférico. En cambio, cuando el simbionte sea común el huésped escogerá la vía rápida: capturarlo (transmisión horizontal).
Entre ambas posibilidades existen niveles de integración que a menudo se interpretan como fases intermedias donde un endosimbionte evoluciona hacia un orgánulo. Pero existen 2 criterios que todo orgánulo celular debe cumplir: 1) estar integrado en la arquitectura celular del huésped y dividirse de forma sincronizada con él, y 2) sobrevivir gracias a proteínas codificadas por el núcleo del huésped porque han perdido la capacidad de existir en la naturaleza como forma libre.
La prueba de ambos criterios es lo que han presentado Coale y col. en su último trabajo en Science. En la portada no veréis dodecaedros porque estudiaron al flagelado. Y las bolitas naranjas son las cianobacterias o mejor dicho: los nitroplastos.

Hasta ahora solo se conocían 3 orgánulos procedentes de endosimbiontes: mitocondrias, cloroplastos y cromatóforos. Los dos últimos hacen la fotosíntesis y en su origen fueron cianobacterias. Los cromatóforos solo se han encontrado en Paulinella chromatophora, una microalga de agua dulce y salobre…

Paulinella chromatophora. Fuente: plingfactory.de
El nitroplasto también procede de cianobacterias. Esta palabra no es nueva. Se usó por primera vez en 1994 para referirse a las cianobacterias simbiontes de diatomeas como posibles orgánulos. No había pruebas genéticas y metabólicas que apoyasen aquella idea pero en el caso de B. bigelowi/C. parkeae ya las tenemos.
El simbionte es único en cada célula de B. bigelowii. Está rodeado por una membrana interna del huésped cubierta por mitocondrias que deben suministrarle energía. Se divide sincronizadamente con el huésped lo cual asegura su transmisión a la descendencia: primero se divide el simbionte, después el núcleo de B. bigelowii cpn sus cloroplastos y por último se separan dos células hijas idénticas.
El nitroplasto importa muchas proteínas fabricadas por el huésped y este las etiqueta para que lleguen a buen puerto usando un sistema análogo al de orgánulos como los cloroplastos. Sin esta ayuda el nitroplasto no tendría ritmos de división; ni siquiera sobreviviría porque no fija carbono, ni hace fotosíntesis ni puede fabricar algunas moléculas esenciales para la vida.
Su única función conocida es la de fijar nitrógeno para B. bigelowii y este le entrega a su vez lo necesario para vivir. Su asociación data de aprox. 100 millones de años, similar al tiempo calculado para las cianobacterias endosimbiontes de diatomeas o el cromatóforo de Paulinella. Esto hace del nitroplasto un orgánulo muy reciente comparado a los cloroplastos (1500 millones de años).
No puedo terminar sin citar otro estudio que me llamó mucho la atención. Unos investigadores japoneses publicaron en 2021 que al menos una cepa de B. bigelowii perdió al nitroplasto durante sus experimentos en el laboratorio. Y sobrevivió sin él. De hecho esa cepa (con el código 3865) continúa en el catálogo de la colección NIES.

Fuente: NIES collection
Si volvéis a leer la portada de Science veréis que se titula «más allá de la simbiosis: se acumulan evidencias sobre un orgánulo fijador de nitrógeno» y los propios autores del trabajo, Coale y col., explican que el nitroplasto funciona «como un orgánulo en una fase temprana de evolución«. Averiguar por qué puede ser inestable en el laboratorio es una cuestión intrigante…
Hay otras pruebas del nitroplasto como orgánulo. Y estas llegan de observaciones naturales. En el oceáno existen variedades de B. bigelowii con tamaños muy diferentes. Seguramente se trata de especies cercanas y todas ellas tienen simbiontes «Candidatus…» relacionados entre sí (UCYN-A1, A2 y A3). Pues bien. Los simbiontes A1, A2 y A3 mantienen un tamaño proporcional a su huésped tal y como sucede con otros orgánulos (p.ej. los cloroplastos).

Fuente: Cornejo-Castillo y col (2024).
En resumen, una noche de luna llena en el Cretácico Braarudosphaera bigelowii capturó una cianobacteria. Aquel encuentro evolucionó a una simbiosis con una versión reducida de la cianobacteria: el nitroplasto, que se comporta como un orgánulo celular. En millones de años quizás lo capturen otras microalgas y si lo conservan se expandirán por el árbol de la vida igual que sucedió con mitocondrias y cloroplastos.
Referencias:
- Cabello AM y col. Unexpected presence of the nitrogen-fixing symbiotic cyanobacterium UCYN-A in Monterey Bay, California. J. Phycol. 56:1521–1533 (2020).
- Coale TH y col. Nitrogen-fixing organelle in a marine alga. Science 384:217–222 (2024).
- Cornejo-Castillo FM y col. Metabolic trade-offs constrain the cell size ratio in a nitrogen-fixing symbiosis. Cell 187:1762–1768 (2024).
- Deflandre, G. Braarudosphaera nov. gen., type d’une famille nouvelle de Coccolithophorides actuels a elements composites. Comp. Hebdomadaires Séances l’Acad Sci Paris. 225:439–41 (1947).
- Delaye L y col. How really ancient is Paulinella chromatophora? PLoS Curr. 15:8 (2016).
- Eikrem W y col. Haptophyta. In: Archibald, J., Simpson, A., Slamovits, C. (eds) Handbook of the Protists. Springer, 61 pp. (2017).
- Gran HH & Braarud T. A quantitative study of the phytoplankton in the Bay of Fundy and the Gulf of Maine (including observations on hydrography, chemistry and turbidity). J. Biol. Board Can. 1:279–467 (1935).
- Green JC & Leadbeater BSC. Chrysochromulina parkeae sp. nov. (Haptophyceae) a new species recorded from S. W. England and Norway. JMBA UK 52: 469-474 (1972).
- Gruber A. What’s in a name? How organelles of endosymbiotic origin can be distinguished from endosymbionts. Microbial Cell 6(2): 134-141 (2019).
- Hagino K y col. Discovery of an endosymbiotic nitrogen-fixing cyanobacterium UCYN-A in Braarudosphaera bigelowii (Prymnesiophyceae). PLoS ONE 8:e81749 (2013).
- Jones HL y col. Size and shape variation in the calcareous nannoplankton genus Braarudosphaera following the Cretaceous/Paleogene (K/Pg) mass extinction: clues as to its evolutionary success. Paleobiology 47(4): 680–703 (2021).
- Massana R. The nitroplast: A nitrogen-fixing organelle. Science 384:160-161 (2024).
- Moisander PH y col. Unicellular cyanobacterial distributions broaden the oceanic N2 fixation domain. Science 327:1512–4 (2010).
- Sagan L. On the origin of mitosing cells. J Theor Biol. 1967;14:225–274.
- Schouten S y col. Endosymbiotic heterocystous cyanobacteria synthesize different heterocyst glycolipids than free-living heterocystous cyanobacteria. Phytochem. 85:115-121 (2013).
- Suzuki S y col. Unstable relationship between Braarudosphaera bigelowii (= Chrysochromulina parkeae) and its nitrogen-fixing endosymbiont. Front. Plant Sci. 12:749895 (2021).
- Thompson A y col. Unicellular cyanobacterium symbiotic with a single-celled eukaryotic alga. Science 337:1546–50 (2012).
- Wouters J y col. The luggage hypothesis: Comparisons of two phototrophic hosts with nitrogen-fixing cyanobacteria and implications for analogous life strategies for kleptoplastids/secondary symbiosis in dinoflagellates. Symbiosis 49:61-70 (2009).
- Zehr JP y col. Unicellular cyanobacteria fix N2 in the subtropical north Pacific Ocean. Nature 412:635–8 (2001).
- Zehr JP y col. Globally distributed uncultivated oceanic N2-fixing cyanobacteria lack oxygenic photosystem II. Science 322:1110–2 (2008).