
Todo a la vez ¿en todas partes?
Imagen de portada: Michelle Yeoh – «Todo a la vez en todas partes» [Dan Kwan, Daniel Scheinert, 2022]. Fuente: filmaffinity
Todo a la vez en todas partes es una película caótica y excesiva en la que cuesta entrar, pero para mí merece la pena (sin duda) por su segunda mitad.
A partir de aquí os aviso de un pequeño spóiler para enlazar esta cinta con la entrada de hoy. Si no queréis que os adelante nada del argumento saltaos el próximo párrafo y continuad desde FIN DEL SPOILER.
…La protagonista, Evelyn (Michelle Yeoh), descubre la existencia de múltiples universos paralelos con distintas versiones de si misma y que (por razones que no vienen al caso) explotará para sobrevivir en una nueva realidad donde todo es impredecible y muda constantemente. Evelyn triunfaba en ciertos universos pero no en su realidad inicial hasta que consigue dominar las conexiones multidimensionales.
FIN DEL SPOILER
La idea de que todo está en todas partes es la primera parte de un dogma sobre los microorganismos marinos (fitoplancton incluido por supuesto), que propuso el científico holandés Baas Becking en 1934 y cuyo enunciado completo es todo está en todas partes, pero el ambiente selecciona. Este simple y bello postulado ha inspirado numerosos estudios de biodiversidad y biogeografía. Viene a decir que el mar no tiene barreras y que por ello procariotas y protistas están en todas partes, pero solo abundan bajo condiciones favorables.

Lourens Baas Becking (1895-1963). Fuente: Fotógrafo desconocido; Wikimedia (Fotocollectie Dienst voor Legercontacten Indonesië – GaHetNa (Nationaal Archief NL) Bestanddeelnr 98-2-5, CC0).
¿Esto es realmente cierto? ¿y a qué escala?
Hutchinson definía en 1957 el nicho ecológico de la población de una especie como el espacio multidimensional donde cada dimensión representa la respuesta a cada variable. Dentro de su nicho la especie puede persistir indefinidamente. El fitoplancton conquista el mar todos los días pero si bajamos al detalle de los organismos que lo forman, las capacidades de crecimiento y adaptación de sus individuos son limitadas.
El mar tiene barreras. Y estas (como la luz, temperatura, turbulencia, salinidad, nutrientes, predadores, corrientes…) se bastan para aislar regiones enteras, generar eventos de especiación a lo largo del tiempo e impedir de facto que todas las especies estén en todas partes.
Las poblaciones costeras y de océano abierto son diferentes y dentro de una misma región biogeográfica (p.ej. un mar interior o el Atlántico Norte) encontramos poblaciones distintas cuyo aislamiento no se puede entender sin esas barreras que delimitan (como muros de ladrillo) la distribución de las especies. El espacio multidimensional de cada especie, por amplio que sea, no supera a la estructura que impone el medio marino.
Por ejemplo, un estudio en el mar Egeo identificó los grupos y géneros de fitoplancton mediante microscopía y genética en 9 estaciones distribuidas en dicho mar. Pues bien, Spatharis y col. (2019) descubrieron que las poblaciones de dichas estaciones eran más distintas entre sí de lo que cabría esperar si solo fuese el ambiente el que selecciona a los organismos.
Cada estación mostraba diferencias según las condiciones locales, pero había una brusca división en 2 cuencas marinas (este y oeste) por la circulación de masas de agua hacia el norte en el este y hacia el sur en el oeste (donde se mezclan aguas salobres del mar Negro antes de emprender el regreso hacia el Mediterráneo). Casi un tercio de los géneros de fitoplancton no aparecían en ambas regiones y algunos muy abundantes en el oeste estaban ausentes en el este.
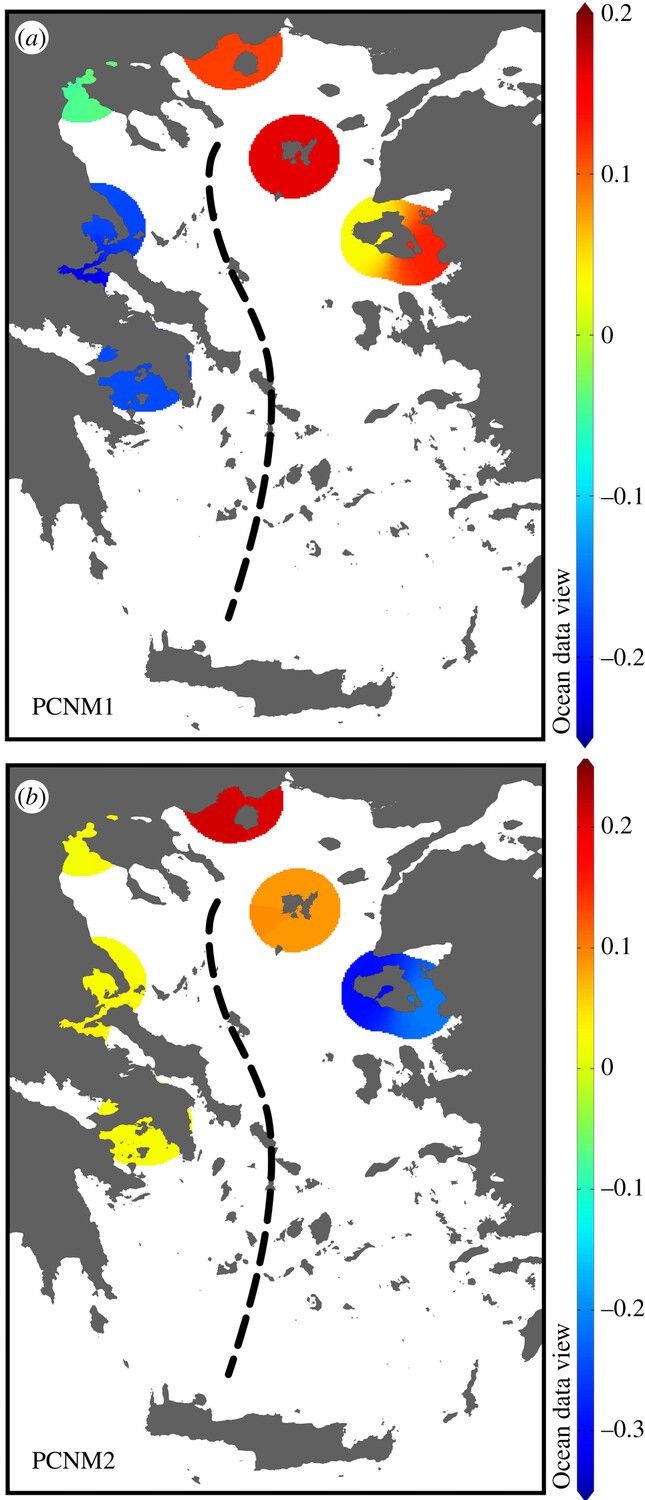
Variación espacial de coordenadas principales (PCNM: matrices de vecinos), un método de análisis que sirve para crear variables que capturan patrones espaciales. Las coordenadas 1 y 2 explican la mayor parte de variabilidad del fitoplancton en las 9 estaciones (agrupadas por colores según sus patrones en dichas coordenadas) y definen la separación entre el este y oeste del Egeo. Fuente: Spatharis y col. (2019).
En muchos trabajos podemos leer que «el grupo XXX» o «el género YYY» de microalgas tienen distribución global, pero nunca que una especie tenga distribución global. En otras ocasiones, lo que parecía una especie global se ha descubierto que incluía varias especies crípticas (que con una nueva técnica se consiguen diferenciar) >>>> El organismo fotosintético más abundante, la cianobacteria Prochlorococcus, posee solo una especie formalmente descrita (P. marinus) pero reconocemos dos subespecies (ecotipos) aclimatadas a luz baja y alta (P. marinus subsp. marinus y P. subsp. pastoris) con múltiples subecotipos que extienden su distribución entre 40ºN y 40ºS. Sin embargo, los estudios genéticos más recientes apoyan que esas variantes equivaldrían a múltiples especies de Prochlorococcus (Dvořák y col. 2023).
Prochlorococcus marinus. Autores: Luke Thompson (Chisholm Lab) y Nikki Watson (Whitehead, MIT). Fuente:Wikimedia.
Las especies de fitoplancton no están en todas partes (ni falta que les hace). La vida se abre paso con la navaja multiuso de la diversidad para colonizar todos los espacios posibles. Pero a nivel de especie esto no sucede. Su navaja es limitada pero las especies no son «ideales estáticos» y su propia variabilidad (importante en microorganismos como el fitoplancton) facilita su adaptación al medio.
Como ejemplo, Brandenburg y col (2018) estudiaron 15 cepas del dinoflagelado Alexandrium ostenfeldii aisladas (a) del mar Báltico y (b) de un arroyo costero (el Ouwerkerkse). Pues bien. Encontraron diferencias de hasta 3 veces en el crecimiento, 5 veces en su biovolumen…¡¡y 81 veces en su contenido de toxinas!!, por citar algunas de las 15 variables que analizaron. Las diferencias eran enormes entre cepas del mismo origen y también comparando las poblaciones. En este caso, las cepas Bálticas crecían más rápido, eran más pequeñas y escasamente tóxicas. Por su parte, las del arroyo impronunciable eran mayores, de crecimiento lento y muuuuy tóxicas. También tenían menor afinidad por los nutrientes, como si nadaran en la abundancia.
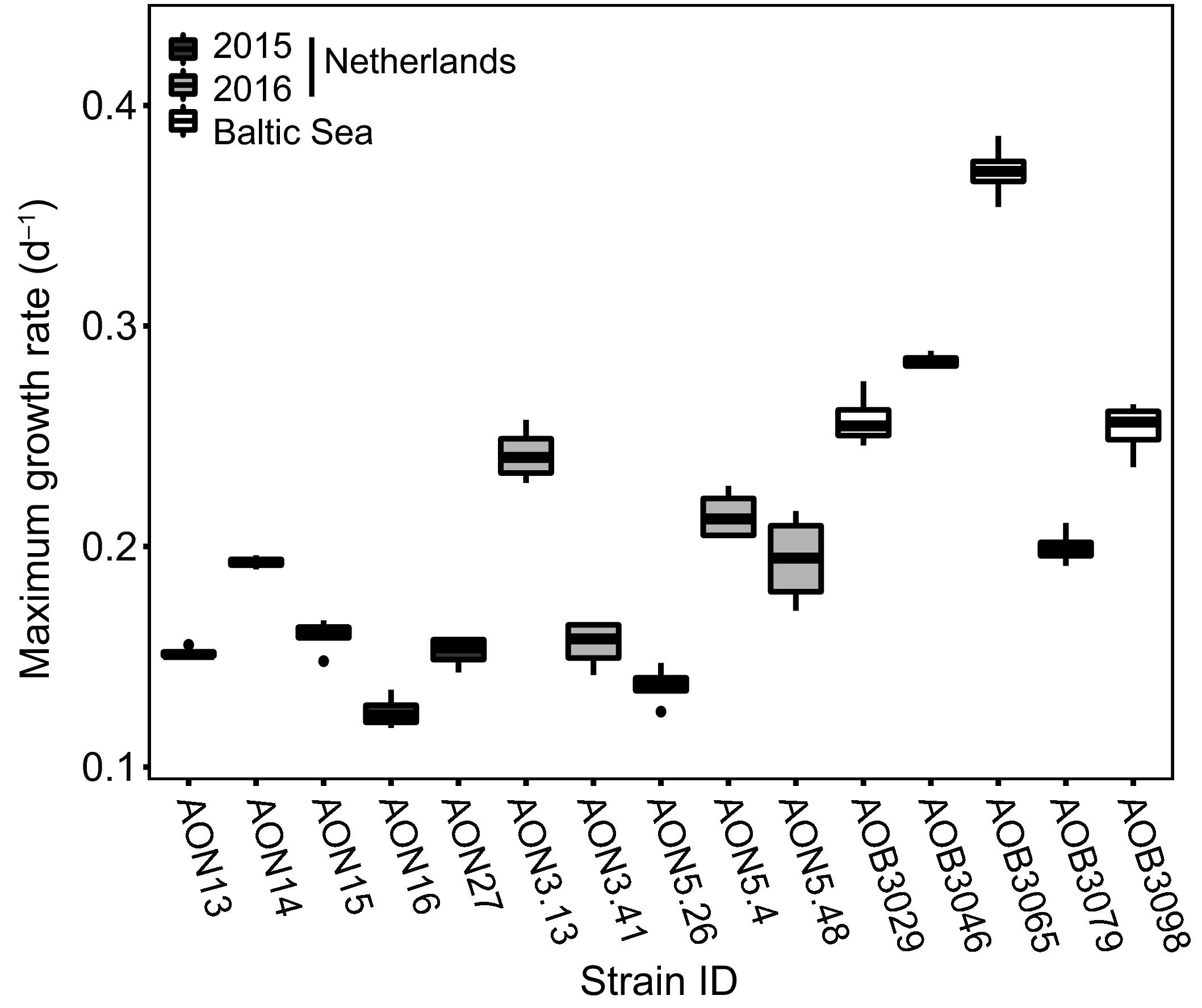
Diferencias en las tasas de crecimiento de las 15 cepas de Alexandrium ostenfeldii aisladas en el Báltico y el arroyo Ouwerkerkse en 2015 y 2016. Fuente: Brandenburg y col. (2018). Las de la derecha son las del Báltico, que crecen como motos…
Las cepas más tóxicas desvían mayor energía hacia su producción por lo que crecen menos y su tamaño aumenta. Esto es así porque en el fitoplancton cuanto más pequeño más rápido creces (y viceversa). La versión supertóxica que desarrolla A. ostenfeldii en el arroyo es una adaptación a las condiciones ambientales. El arroyo posee más nutrientes que el Báltico, inestabilidad ambiental, mayor competencia con otras microalgas y presión de predadores. En este contexto la toxicidad es ventajosa como defensa frente al zooplancton y atacando a otras microalgas con sustancias alelopáticas expulsadas al medio. Crecer menos y ser más grande es consecuencia de dicha estrategia, pero todo compensa.
Alexandrium ostenfeldii (cepa AOSH1, IMB culture collection). Autor: National Research Council of Canada, Nancy Lewis. Fuente: marinespecies.org
El secreto que ha permitido persistir a la vida en nuestro planeta no son las especies individuales sino la plasticidad de los individuos en sus poblaciones.
Las versiones que cada especie crea de sí misma son sus armas para competir en un mundo cambiante…tal y como le sucede a Evelyn en «Todo a la vez en todas partes».
Referencias
- Brandenburg K.M. y col. Intraspecific trait variation and trade-offs within and across populations of a toxic dinoflagellate. Ecol Lett. 21(10):1561-1571 (2018).
- De Wit R. & Bouvier T. ‘Everything is everywhere, but, the environment selects’; what did Baas Becking and Beijerinck really say? Env. Microbiol. 8:755-758 (2006).
- Dvořák P. y col. Population genomics meets the taxonomy of cyanobacteria. Algal Res. 72:103128 (2023).
- Hutchinson G.E. Concluding remarks. Cold Spring Harbor Symposia on Quantitative Biology. 22:415–427 (1957).
- Spatharis S. y col. Everything is not everywhere: can marine compartments shape phytoplankton assemblages? Proc. R. Soc. B 286: 20191890 (2019).






